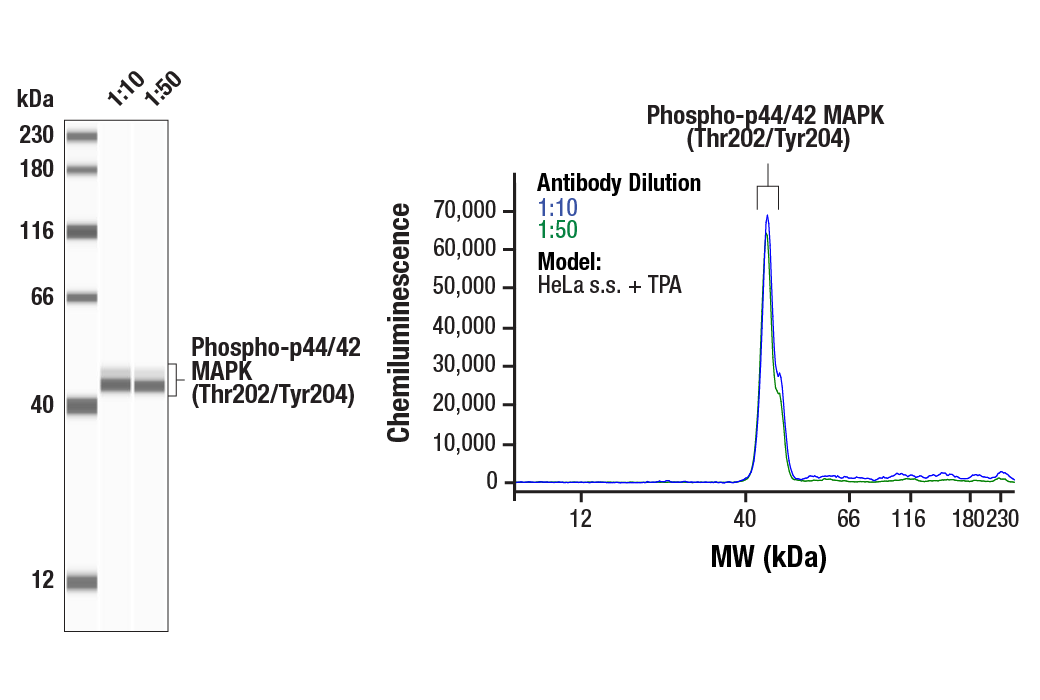
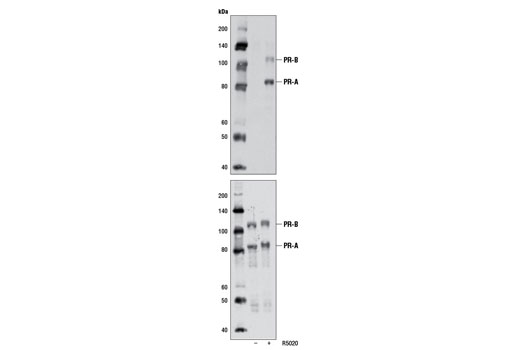
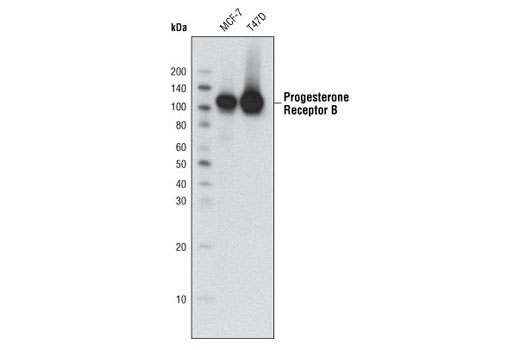
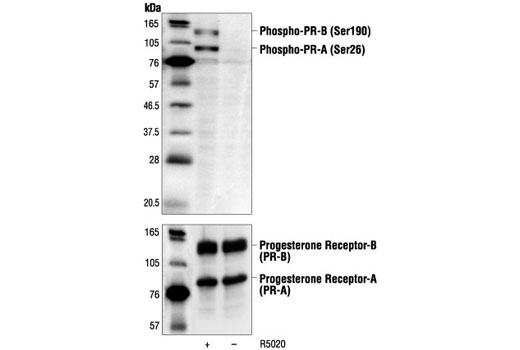
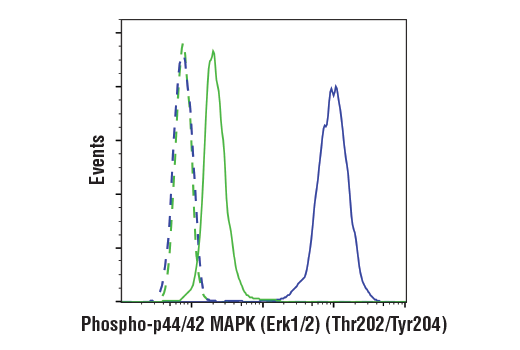
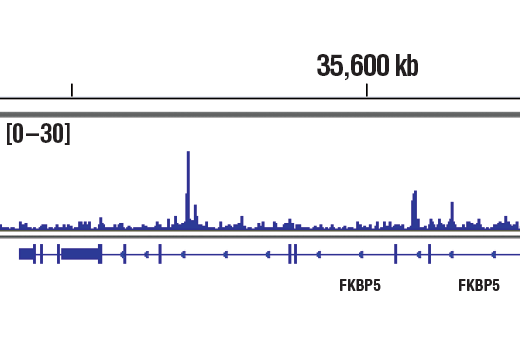
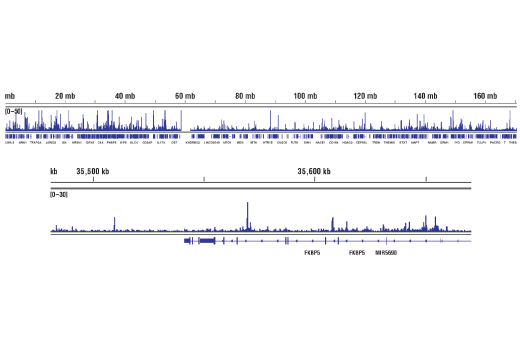
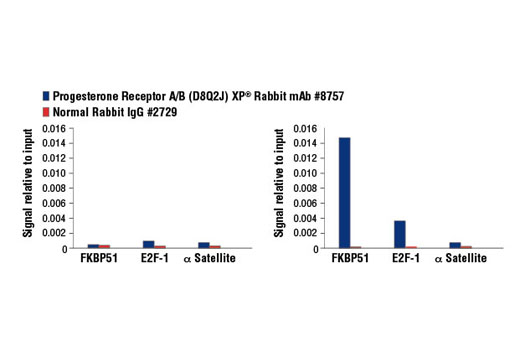
Product Information
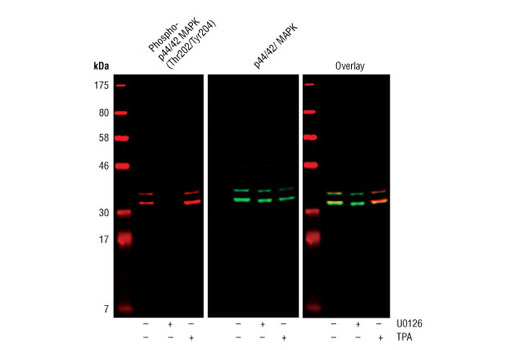
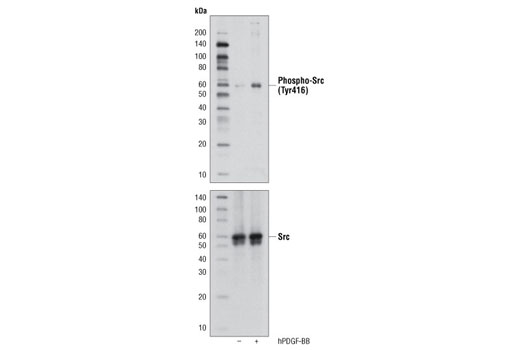
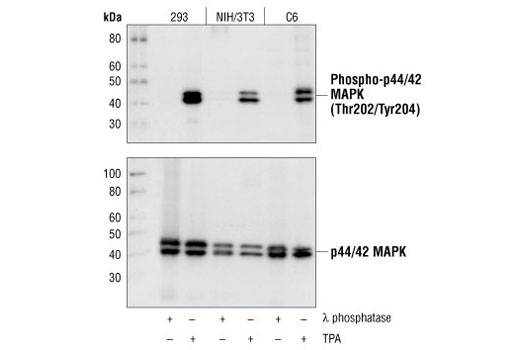
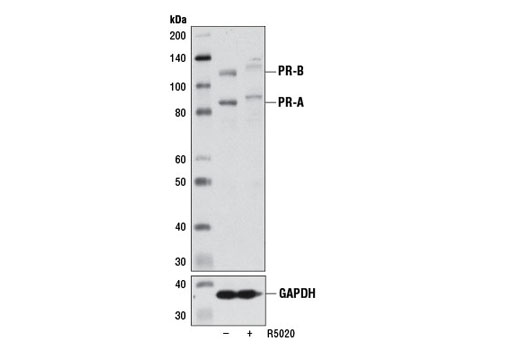
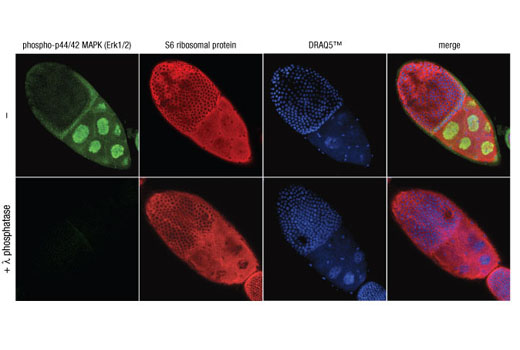
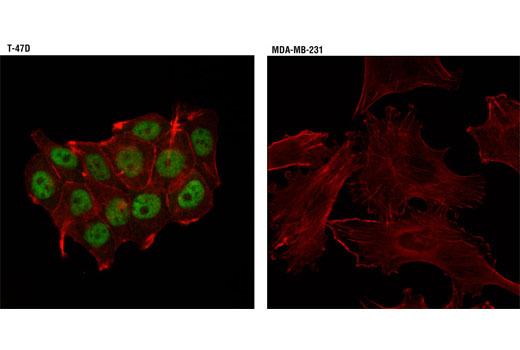
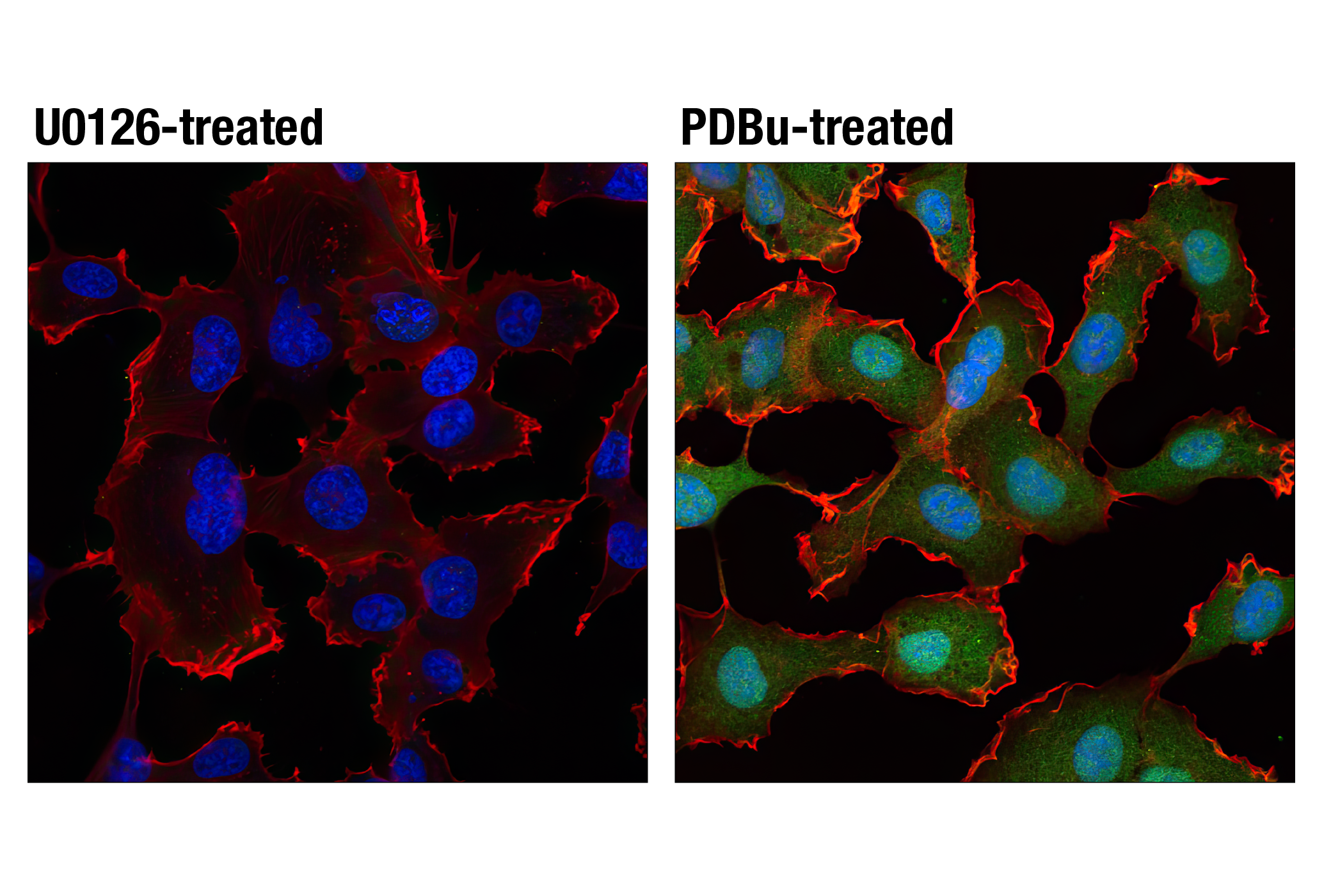
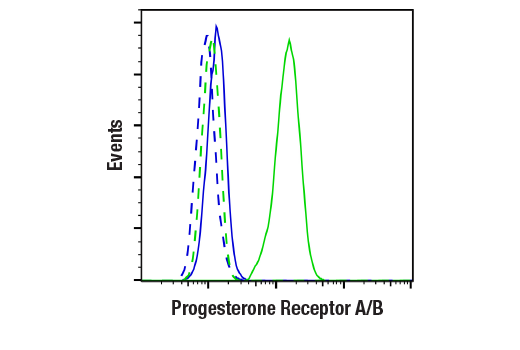
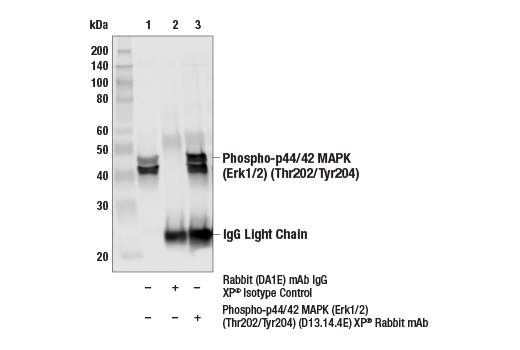
Monoclonal antibodies are produced by immunizing animals with a synthetic phosphopeptide corresponding to residues surrounding Thr202/Tyr204 of human p44 MAP kinase, or with synthetic peptides corresponding to residues surrounding Ser115 or Tyr541 of human progesterone receptor protein, or Tyr416 of human Src protein. Polyclonal antibodies are produced by immunizing animals with synthetic phosphopeptides corresponding to residues surrounding Ser190 of human progesterone receptor protein or Ser345 of human progesterone receptor B protein. Polyclonal antibodies are purified by protein A and peptide affinity chromatography.
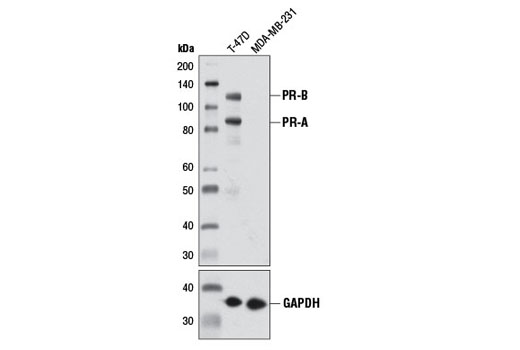
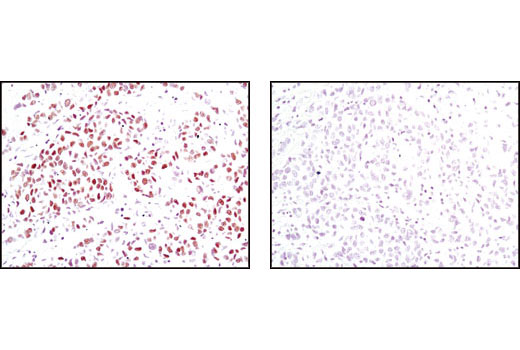
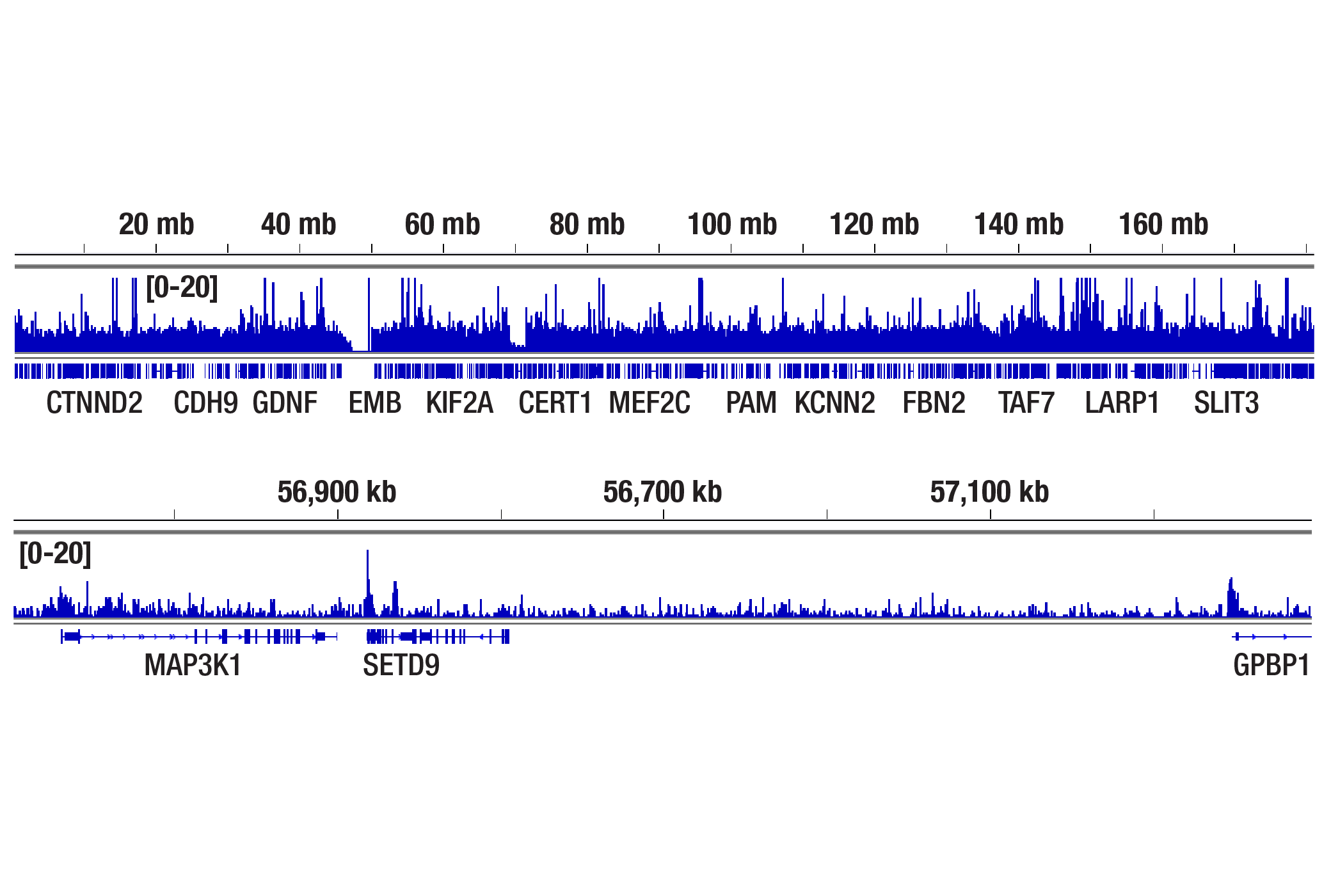
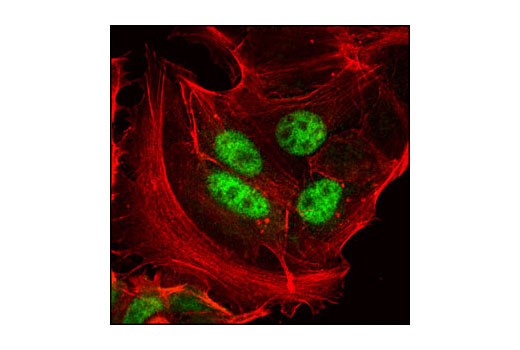
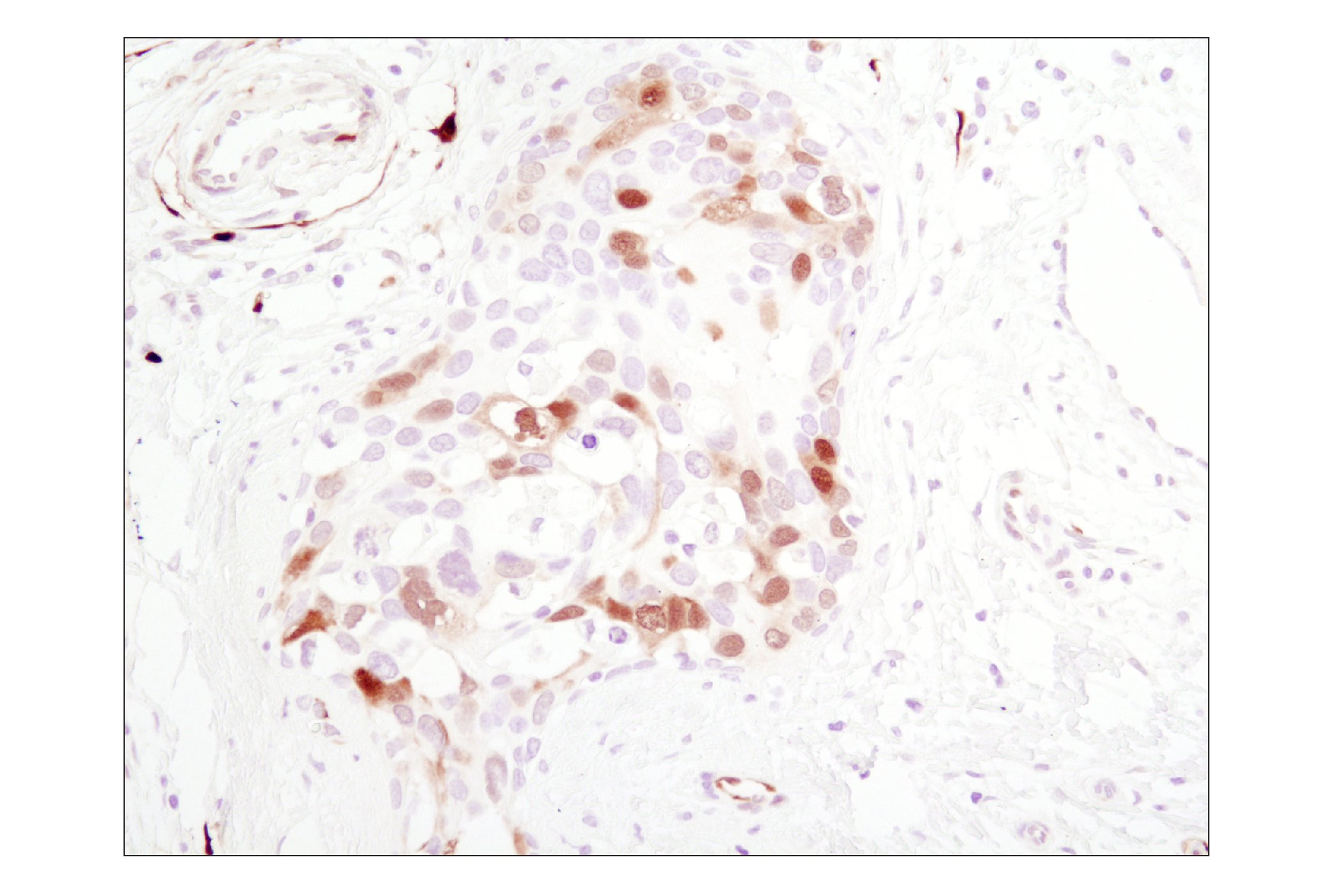
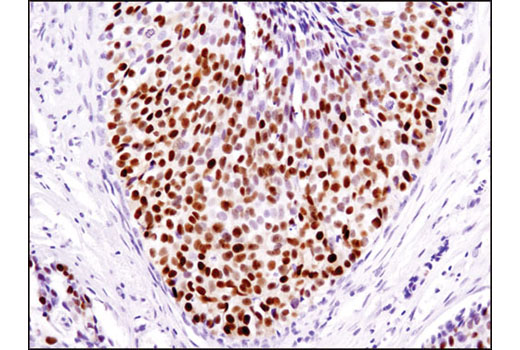
Human progesterone receptor (PR) is expressed as two forms: the full length PR-B and the short form PR-A. PR-A lacks the first 164 amino acid residues of PR-B (1,2). Both PR-A and PR-B are ligand activated, but differ in their relative ability to activate target gene transcription (3,4). The activity of PR is regulated by phosphorylation; at least seven serine residues are phosphorylated in its amino-terminal domain. Three sites (Ser81, Ser102, and Ser162) are unique to full length PR-B, while other sites (Ser190, Ser294, Ser345, and Ser400) are shared by both isoforms (5). Phosphorylation of PR-B at Ser190 (equivalent to Ser26 of PR-A) is catalyzed by CDK2 (6). Mutation of Ser190 results in decreased activity of PR (7), suggesting that the phosphorylation at Ser190 may be critical to its biological function. Research studies have demonstrated ligand-dependent phosphorylation of PR-B at Ser345 is catalyzed by MAPK and plays an important role in mediating the proliferation of breast cancer cells. Investigators have shown that Ser345-phosphorylated PR-B associates with Sp1 to regulate EGFR and p21 transcription (8). PR signaling has been shown to crosstalk with other kinase signaling cascades. Upon stimulation and the subsequent interaction with estrogen receptor α and c-Src, PR-B has been shown to promote the activation of the Src/p21ras/Erk pathway (9).
Explore pathways related to this product.
STRING - Known and Predicted Protein-Protein Interactions.
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.