| Cat. # | Size | Qty. | Price |
|---|---|---|---|
| 8342T | 1 Kit (7 x 20 microliters) |
|
| Product Includes | Quantity | Applications | Reactivity | MW(kDa) | Isotype |
|---|---|---|---|---|---|
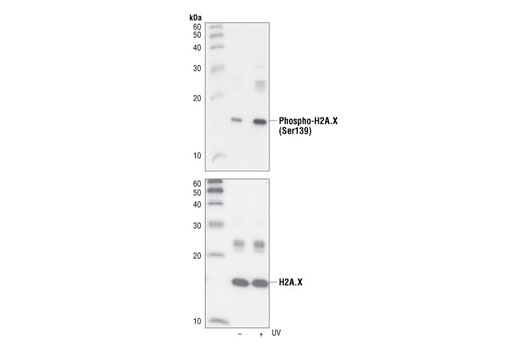
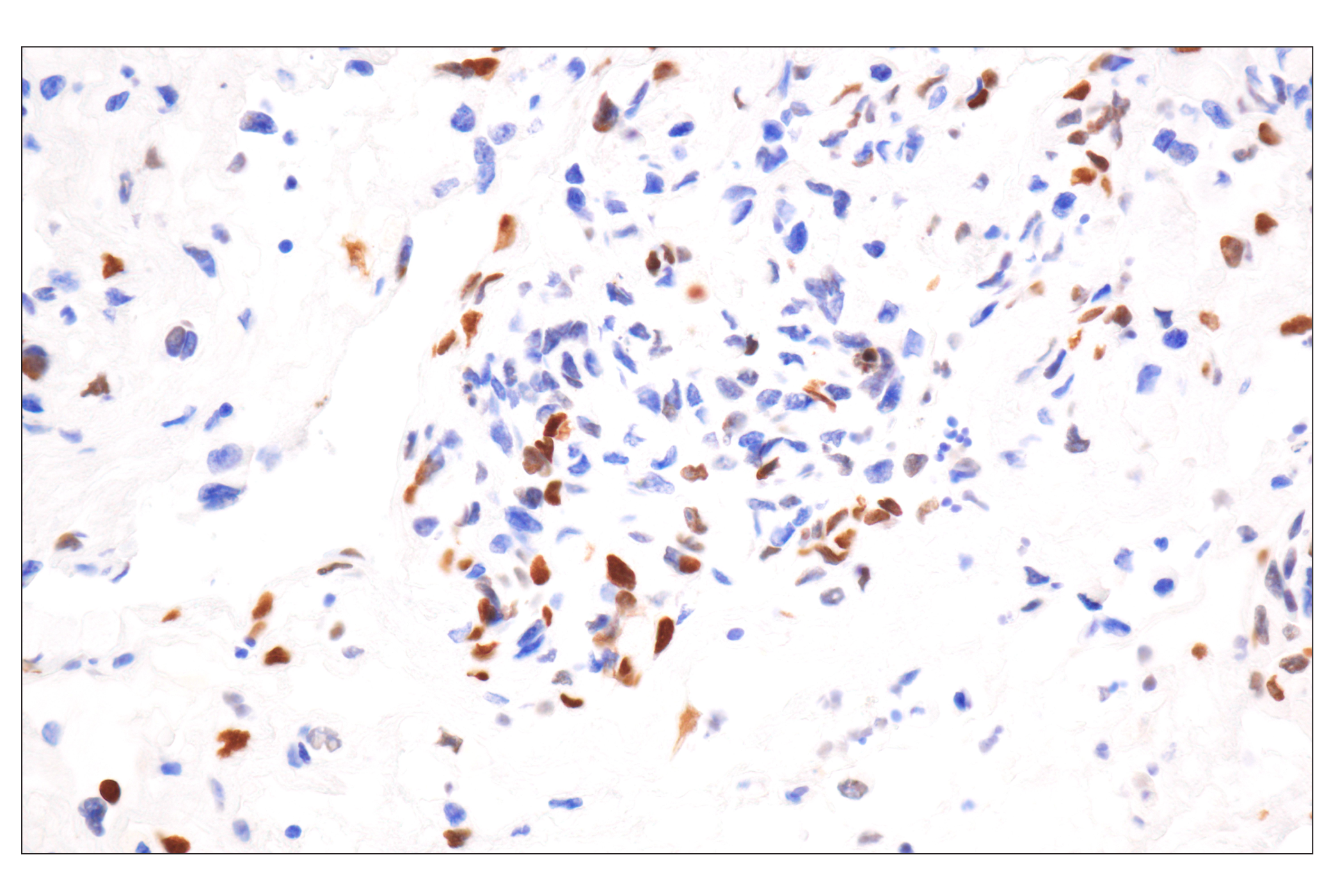
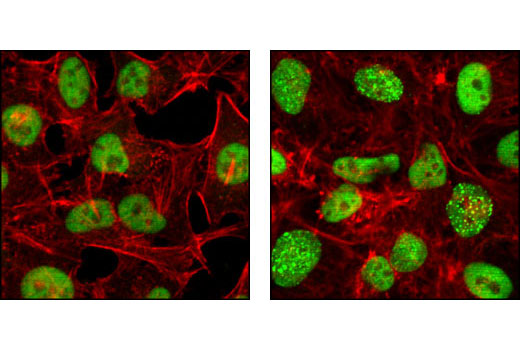
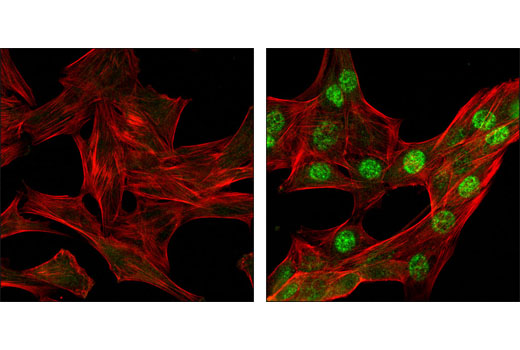
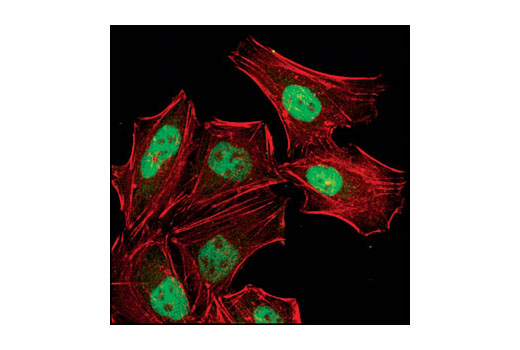
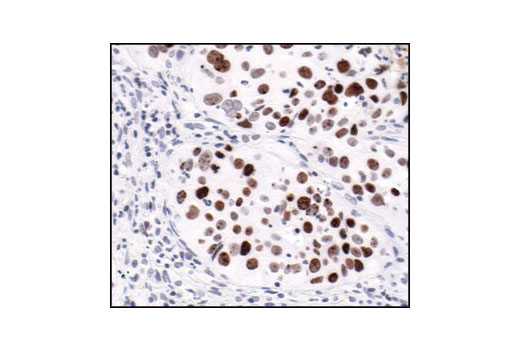
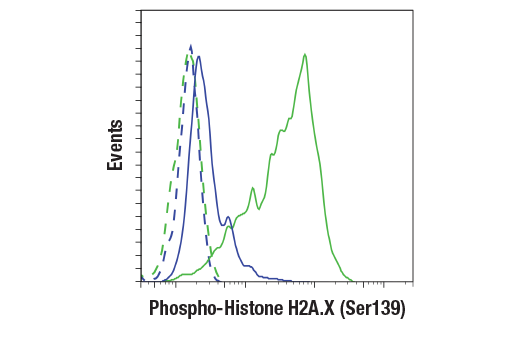
| Phospho-Histone H2A.X (Ser139) (20E3) Rabbit mAb 9718 | 20 µl |
|
H M R Mk | 15 | Rabbit IgG |
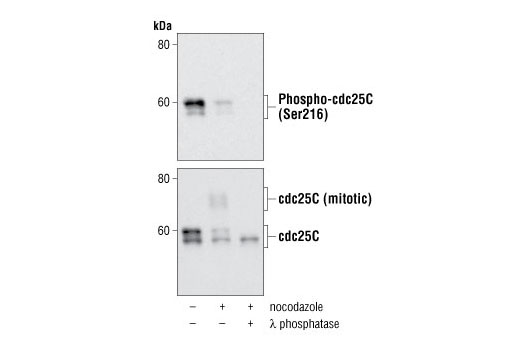
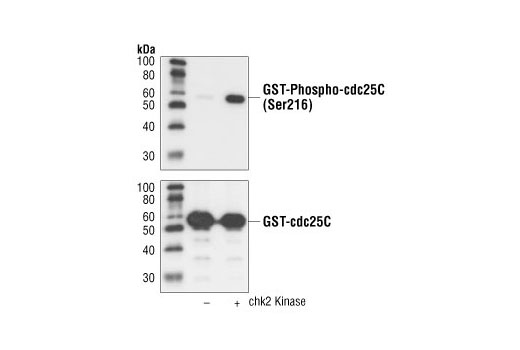
| Phospho-cdc25C (Ser216) (63F9) Rabbit mAb 4901 | 20 µl |
|
H Mk | 60 | Rabbit IgG |
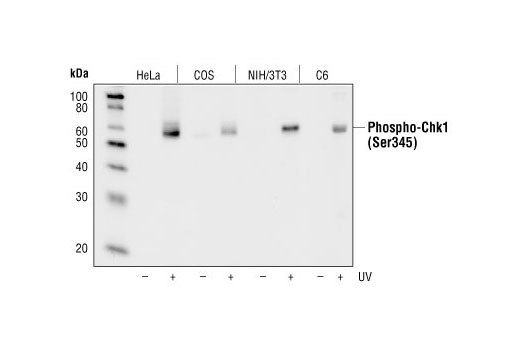
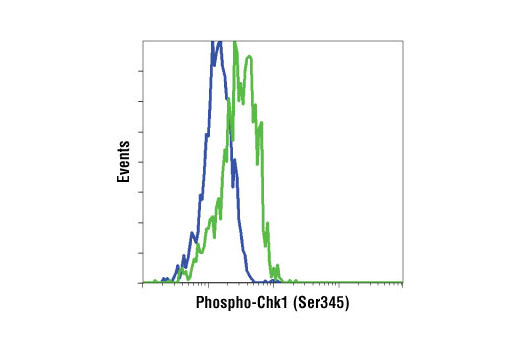
| Phospho-Chk1 (Ser345) (133D3) Rabbit mAb 2348 | 20 µl |
|
H M R Mk | 56 | Rabbit IgG |
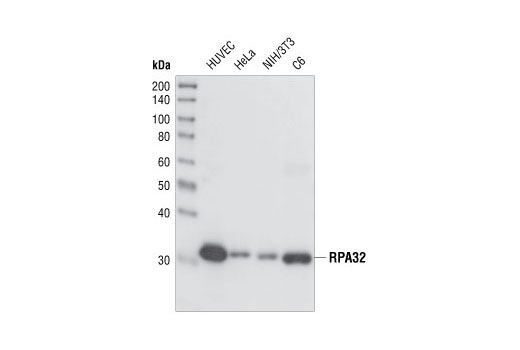
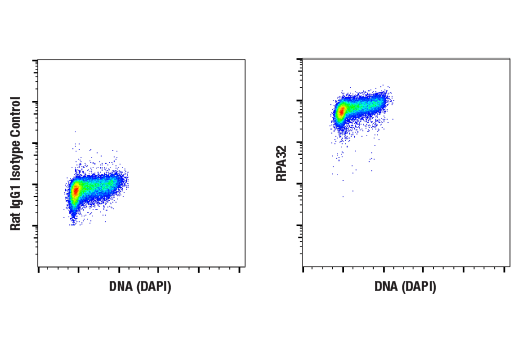
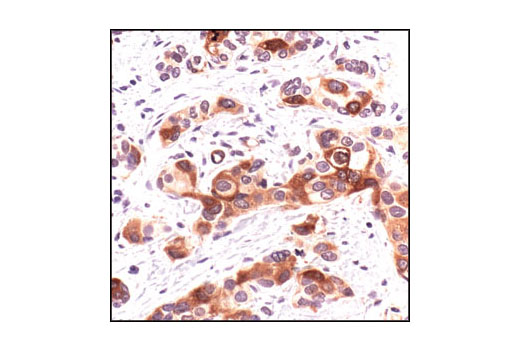
| RPA32/RPA2 (4E4) Rat mAb 2208 | 20 µl |
|
H M R Hm Mk | 32 | Rat IgG1 |
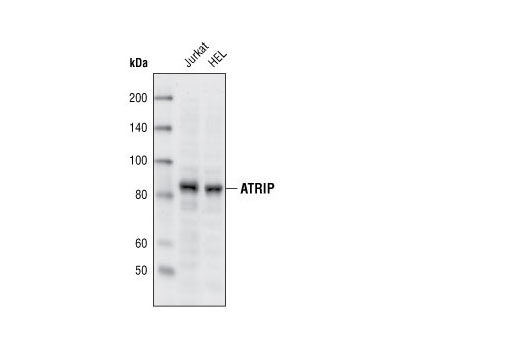
| ATRIP Antibody 2737 | 20 µl |
|
H | 82 | Rabbit |
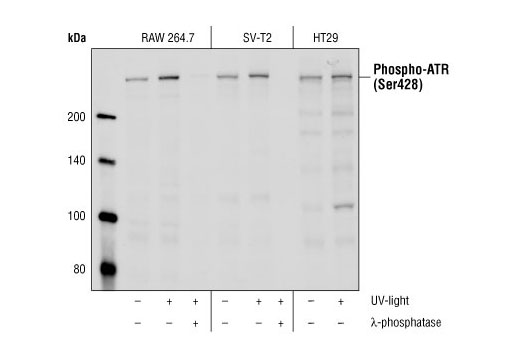
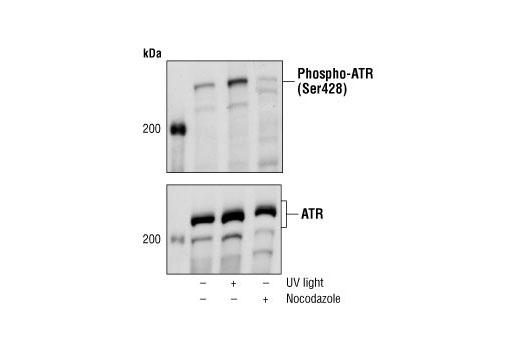
| Phospho-ATR (Ser428) Antibody 2853 | 20 µl |
|
H M R Mk | 300 | Rabbit |
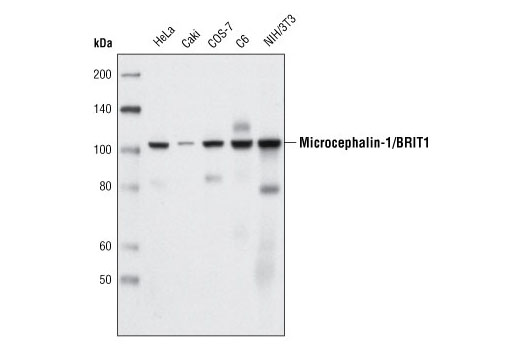
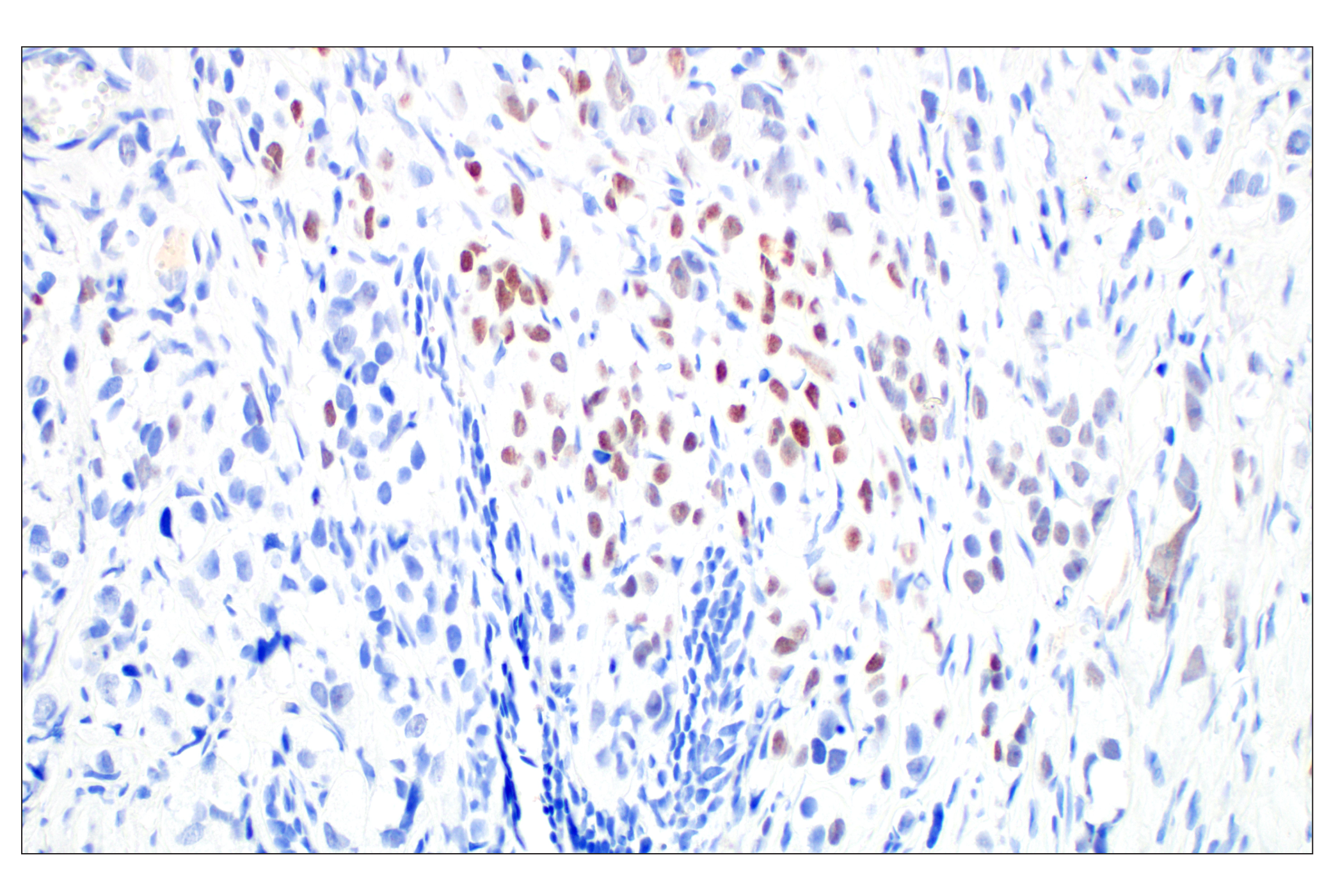
| Microcephalin-1/BRIT1 (D38G5) Rabbit mAb 4120 | 20 µl |
|
H M R Mk | 100 | Rabbit IgG |
| Anti-rabbit IgG, HRP-linked Antibody 7074 | 100 µl |
|
Goat | ||
| Anti-rat IgG, HRP-linked Antibody 7077 | 100 µl |
|
R | Goat |
Product Information
Polyclonal antibodies are produced by immunizing animals with a synthetic peptide corresponding to residues surrounding Gly267 of human ATRIP protein or Ser428 of human ATR protein. Polyclonal antibodies are purified by protein A and peptide affinity chromatography. Monoclonal antibodies are produced by immunizing animals with a recombinant full-length human Maltose Binding Protein-RPA32 fusion protein or a synthetic peptide corresponding to residues surrounding Tyr415 of human Microcephalin-1/BRIT1 protein. Activation state monoclonal antibodies are produced by immunizing animals with a synthetic phosphopeptide corresponding to residues surrounding Ser139 of human H2A.X protein, Ser216 of human cdc25C protein, or Ser345 of human Chk1 protein, respectively.
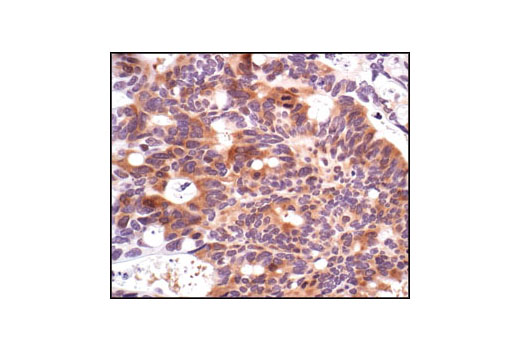
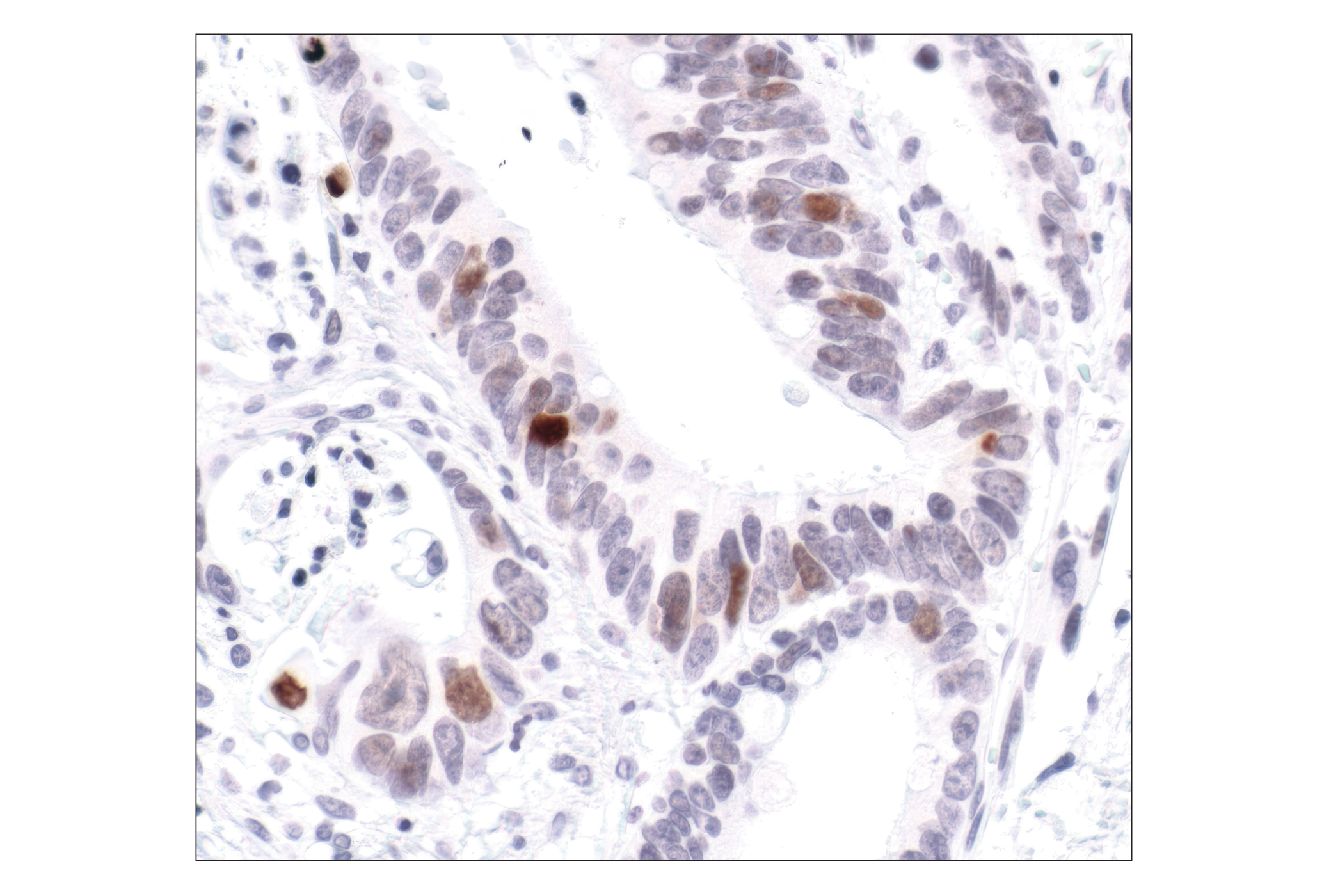
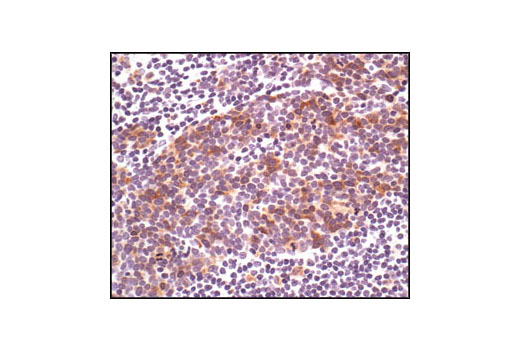
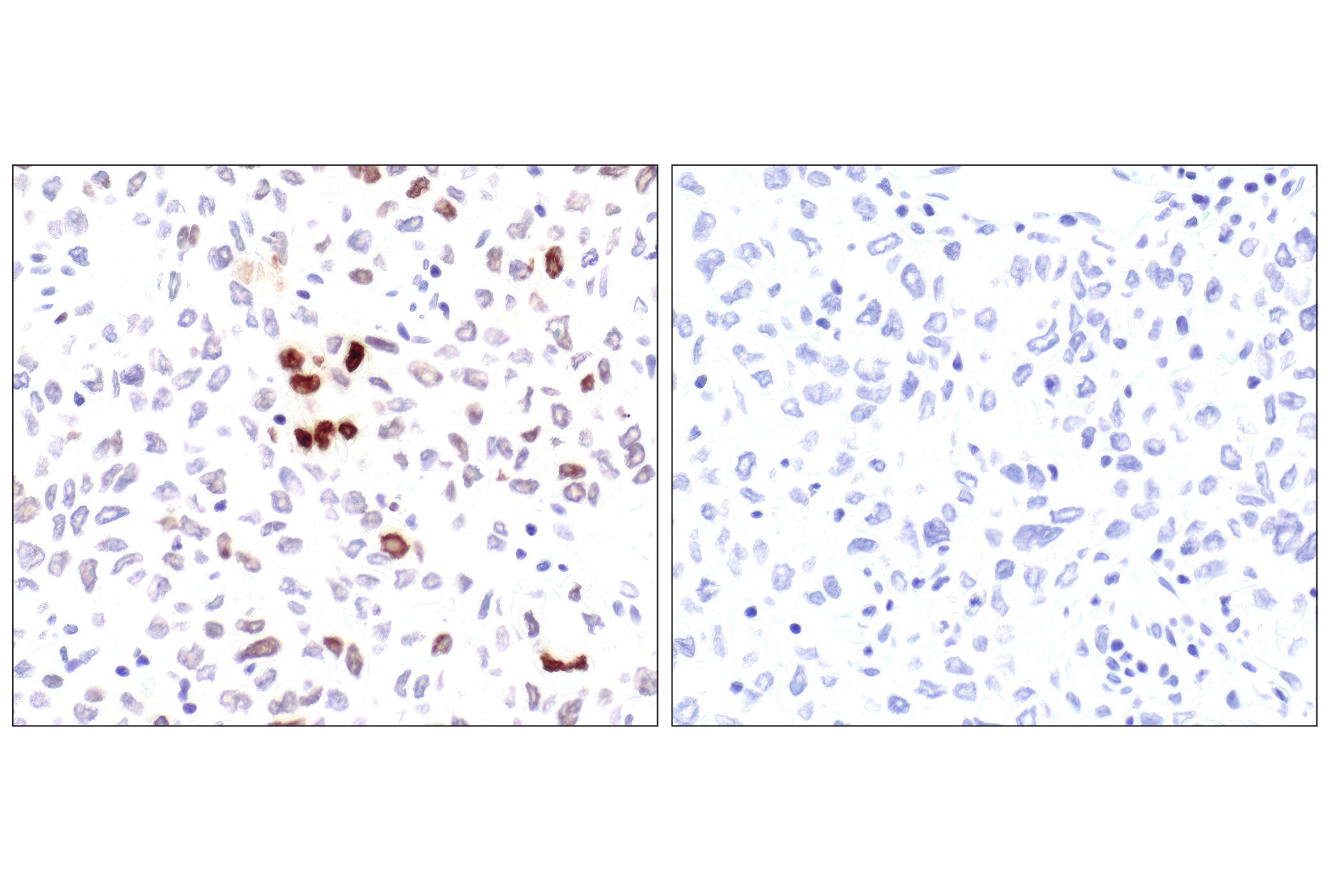
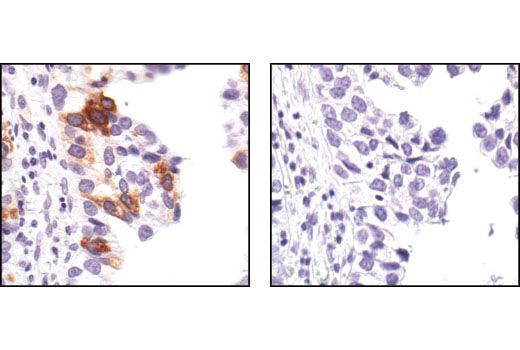
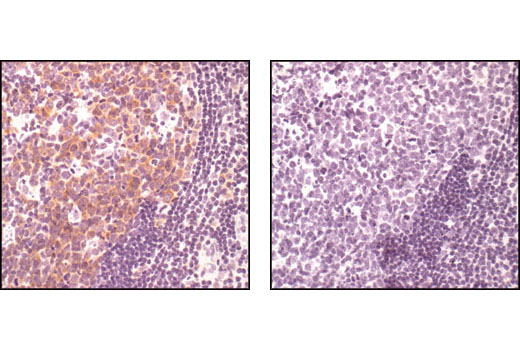
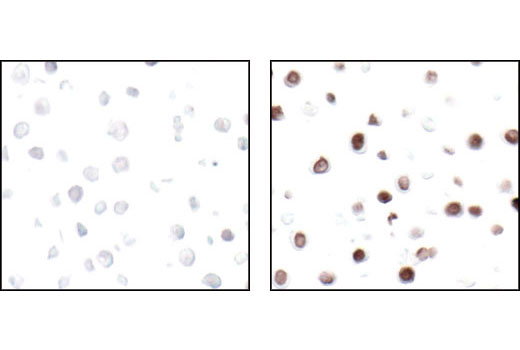
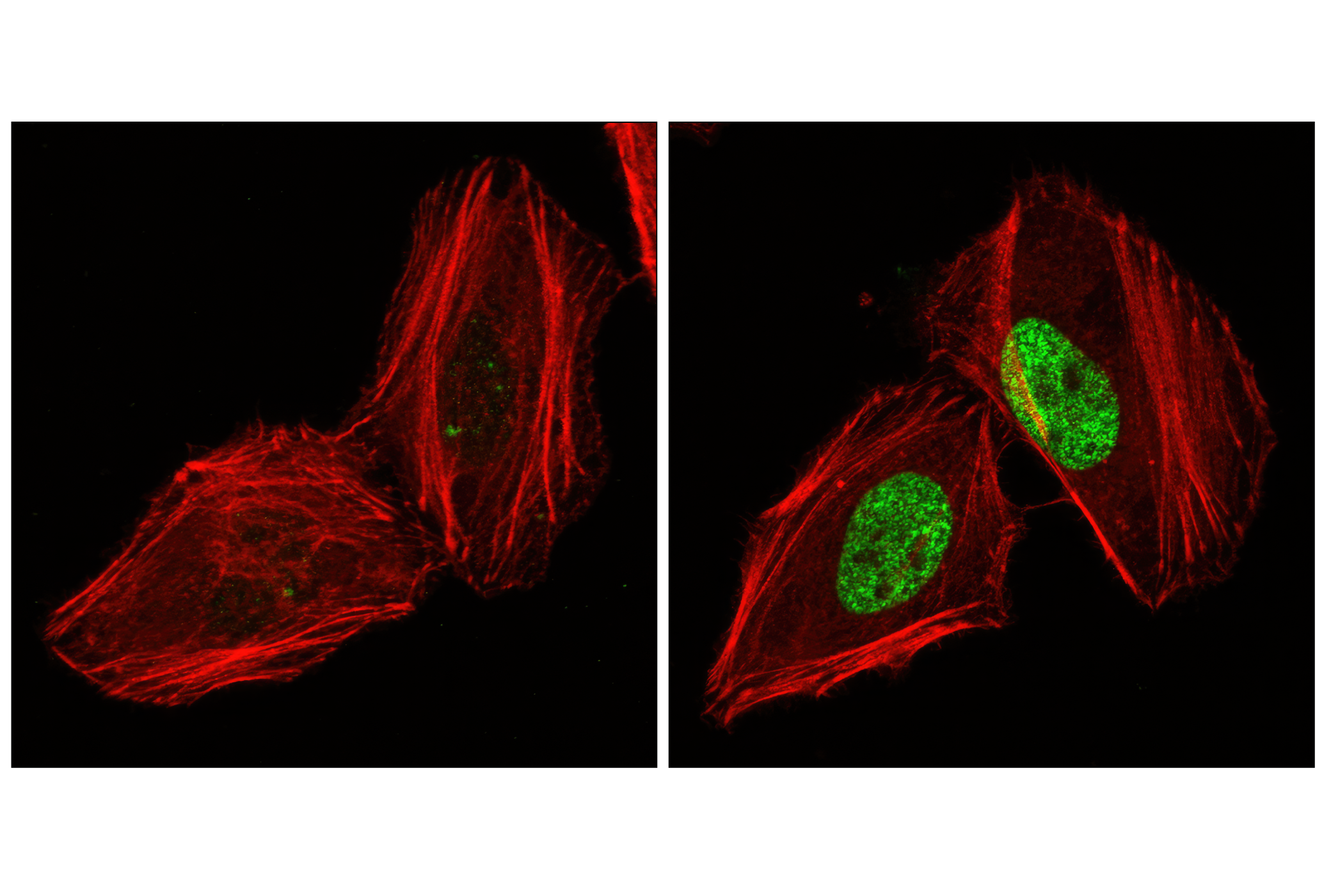
Exposure to ultraviolet radiation (UV) has a profound impact on human health and disease (1). Low level UV exposure induces the production of vitamin D and is a key regulator of calcium metabolism. Conversely, overexposure to UV is associated with an increased risk of cancer, immunosuppression, and many eye disorders, such as cataracts. Photons of UV light can directly damage DNA causing thymine dimers and other pyrimidine dimers between adjacent bases (2). Free radicals and reactive oxygen species induced by UV exposure also result in DNA lesions and have been linked to malignant melanoma (3). DNA damage from replicative stress and genotoxic agents like UV activate the ATR-mediated checkpoint pathway and stimulate DNA repair, cell cycle arrest, and apoptosis (4). ATR recruitment to sites of DNA damage and activation depends, at least in part, on interaction with the complex of single-stranded DNA, Replication Protein A (RPA), and direct binding to the ATR-associated adapter protein, ATRIP (5). In addition, the Rad17-RFC and Rad9-Rad1-Hus1 (9-1-1) protein complexes are independently recruited with TopBP1 to fully activate the checkpoint response (6,7). BRIT1 (MCPH1) is required for UV-induced formation of ATR, RPA, and p-Rad17 foci at sites of DNA damage (8-10) and may regulate the expression of several DNA damage response proteins (11). Once activated, ATR phosphorylates a number of mediators, including histone H2AX Ser139 and Chk1 kinase at Ser345. H2AX phosphorylation is a marker of DNA damage. Complete loss of H2AX results in reduced Chk1 activation and impaired survival of cells after UV exposure (12). Chk1 and Chk2 kinase activation is essential for checkpoint-mediated control of cell cycle progression (4). Checkpoint kinases stimulate cell cycle arrest by phosphorylation of a group of tyrosine phosphatases known as Cdc25A, Cdc25B, and Cdc25C (13 -15). Both Chk1 and Chk2 kinases phosphorylate Cdc25C at Ser216 in response to DNA damage and stimulate arrest (16-17).
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.