| Cat. # | Size | Qty. | Price |
|---|---|---|---|
| 8575T | 1 Kit (6 x 20 microliters) |
|
| Product Includes | Quantity | Applications | Reactivity | MW(kDa) | Isotype |
|---|---|---|---|---|---|
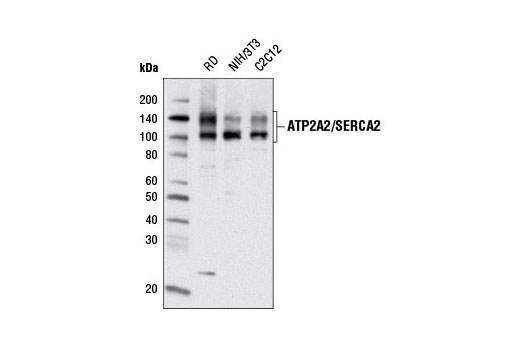
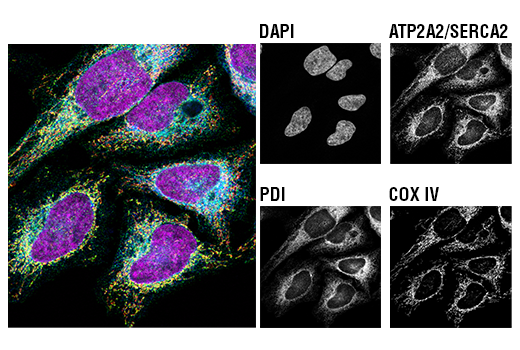
| ATP2A2/SERCA2 (D51B11) Rabbit mAb 9580 | 20 µl |
|
H M R Mk | 114, 140 | Rabbit IgG |
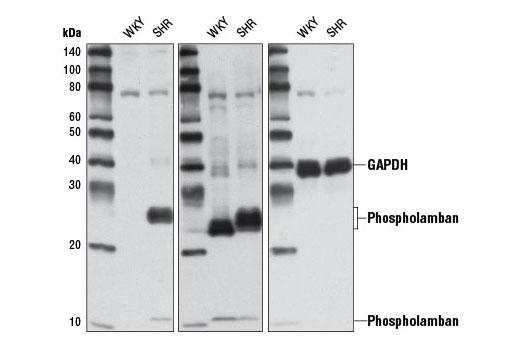
| Phospho-Phospholamban (Ser16/Thr17) Antibody 8496 | 20 µl |
|
R | 6 (monomer); 12, 24 (oligomers) | Rabbit |
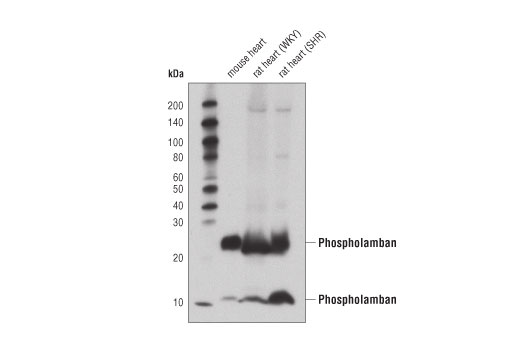
| Phospholamban (D9W8M) Rabbit mAb 14562 | 20 µl |
|
H M R | 12, 24 | Rabbit IgG |
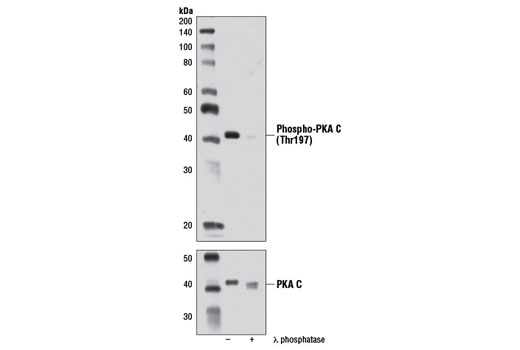
| Phospho-PKA C (Thr197) (D45D3) Rabbit mAb 5661 | 20 µl |
|
H M R Mk | 42 | Rabbit IgG |
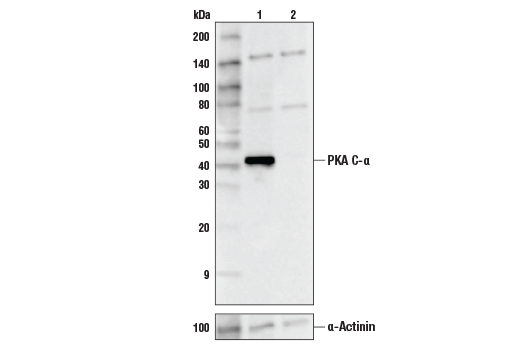
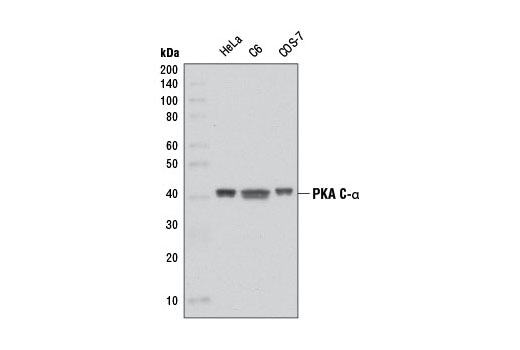
| PKA C-α (D38C6) Rabbit mAb 5842 | 20 µl |
|
H M R Hm Mk | 42 | Rabbit IgG |
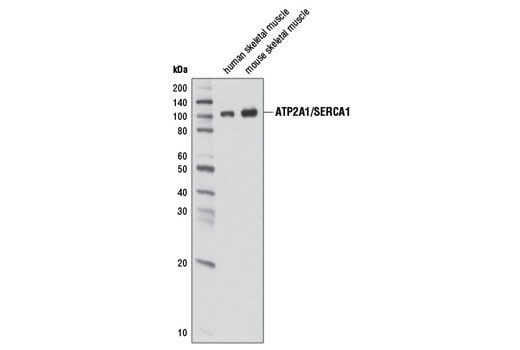
| ATP2A1/SERCA1 (D54G12) Rabbit mAb 12293 | 20 µl |
|
H M | 100 | Rabbit IgG |
| Anti-rabbit IgG, HRP-linked Antibody 7074 | 100 µl |
|
Rab | Goat |
Product Information
Monoclonal antibodies are produced by immunizing animals with a synthetic peptide corresponding to residues near the amino terminus of human ATP2A2/SERCA2 protein, residues surrounding Pro995 of human ATP2A1/SERCA1 protein, residues surrounding Thr197 of human PKA C protein, or residues surrounding Ser326 of human PKA C-α protein. Monoclonal antibodies are also produced by immunizing animals with a synthetic peptide corresponding to residues near the amino terminus of human phospholamban proten (not overlapping Ser16 and Thr17). Phospho-specific polyclonal antibodies are produced by immunizing animals with a synthetic phosphopeptide corresponsing to residues surrounding Ser16/Thr17 of human phospholamban protein. Polyclonal antibodies are purified by protein A and peptide affinity chromatography.
Sarcoplasmic and endoplasmic reticulum Ca2+ ATPases (SERCA) are members of a highly conserved family of Ca2+ pumps (1). ATP2A1 (SERCA1) is a fast-twitch, skeletal muscle sarcoplasmic reticulum (SR) Ca2+ ATPase (2). Multiple ATP2A2 (SERCA2) isoforms have been isolated, with ATP2A2a (SERCA2a) found predominantly in the SR of muscle cells and ATP2A2b (SERCA2b) more ubiquitously expressed in the ER of most cell types (3). Post-translational modification of ATP2A2, including phosphorylation and tyrosine nitration, modify Ca2+ -ATPase activity and calcium transport (4,5).
Phospholamban (PLN) was identified as a major phosphoprotein component of the SR (6). Despite very high expression in cardiac tissue, phospholamban is also expressed in skeletal and smooth muscle (7). Localization of PLN is limited to the SR, where it serves as a regulator of the sarco-endoplasmic reticulum calcium ATPase, SERCA (8). PLN binds directly to SERCA and effectively lowers its affinity for calcium, thus reducing calcium transport into the SR. Phosphorylation of PLN at Ser16 by PKA or myotonic dystrophy protein kinase and/or phosphorylation at Thr17 by Ca2+/calmodulin-dependent protein kinase results in release of PLN from SERCA, relief of this inhibition, and increased calcium uptake by SR (reviewed in 9,10). It has long been held that phosphorylation at Ser16 and Thr17 occurs sequentially, but increasing evidence suggests that phosphorylation, especially at Thr17, may be differentially regulated (reviewed in 11,12).
The second messenger cyclic AMP (cAMP) activates cAMP-dependent protein kinase (PKA or cAPK) in mammalian cells and controls many cellular mechanisms such as gene transcription, ion transport, and protein phosphorylation (13). Inactive PKA is a heterotetramer composed of a regulatory subunit (R) dimer and a catalytic subunit (C) dimer. In this inactive state, the pseudosubstrate sequences on the R subunits block the active sites on the C subunits. Three C subunit isoforms (C-α, C-β, and C-γ) and two families of the regulatory subunits (RI and RII) with distinct cAMP binding properties have been identified. Upon binding of cAMP to the R subunits, the auto-inhibitory contact is eased and active monomeric C subunits are released. PKA shares substrate specificity with Akt (PKB) and PKC, which are characterized by an arginine at position -3 relative to the phosphorylated serine or threonine residue (14). PKA phosphorylation is involved in the regulation of Ca2+ channels, including Cav1.1 in skeletal muscle and Cav1.2 in the heart (reviewed in 15).
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.