| REACTIVITY | H |
Product Information
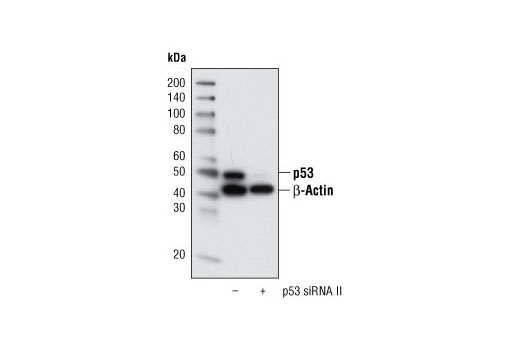
CST recommends transfection with 100 nM p53 siRNA II 48 to 72 hours prior to cell lysis. For transfection procedure, follow protocol provided by the transfection reagent manufacturer. Please feel free to contact CST with any questions on use.
Each vial contains the equivalent of 100 transfections, which corresponds to a final siRNA concentration of 100 nM per transfection in a 24-well plate with a total volume of 300 μl per well.
The p53 tumor suppressor protein plays a major role in cellular response to DNA damage and other genomic aberrations. Activation of p53 can lead to either cell cycle arrest and DNA repair or apoptosis (1). p53 is phosphorylated at multiple sites in vivo and by several different protein kinases in vitro (2,3). DNA damage induces phosphorylation of p53 at Ser15 and Ser20 and leads to a reduced interaction between p53 and its negative regulator, the oncoprotein MDM2 (4). MDM2 inhibits p53 accumulation by targeting it for ubiquitination and proteasomal degradation (5,6). p53 can be phosphorylated by ATM, ATR, and DNA-PK at Ser15 and Ser37. Phosphorylation impairs the ability of MDM2 to bind p53, promoting both the accumulation and activation of p53 in response to DNA damage (4,7). Chk2 and Chk1 can phosphorylate p53 at Ser20, enhancing its tetramerization, stability, and activity (8,9). p53 is phosphorylated at Ser392 in vivo (10,11) and by CAK in vitro (11). Phosphorylation of p53 at Ser392 is increased in human tumors (12) and has been reported to influence the growth suppressor function, DNA binding, and transcriptional activation of p53 (10,13,14). p53 is phosphorylated at Ser6 and Ser9 by CK1δ and CK1ε both in vitro and in vivo (13,15). Phosphorylation of p53 at Ser46 regulates the ability of p53 to induce apoptosis (16). Acetylation of p53 is mediated by p300 and CBP acetyltransferases. Inhibition of deacetylation suppressing MDM2 from recruiting HDAC1 complex by p19 (ARF) stabilizes p53. Acetylation appears to play a positive role in the accumulation of p53 protein in stress response (17). Following DNA damage, human p53 becomes acetylated at Lys382 (Lys379 in mouse) in vivo to enhance p53-DNA binding (18). Deacetylation of p53 occurs through interaction with the SIRT1 protein, a deacetylase that may be involved in cellular aging and the DNA damage response (19).
Explore pathways related to this product.
STRING - Known and Predicted Protein-Protein Interactions.
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.
| Cat. # | Size | Qty. | Price |
|---|---|---|---|
| 6562S | 300 µl (3 nmol) |
|