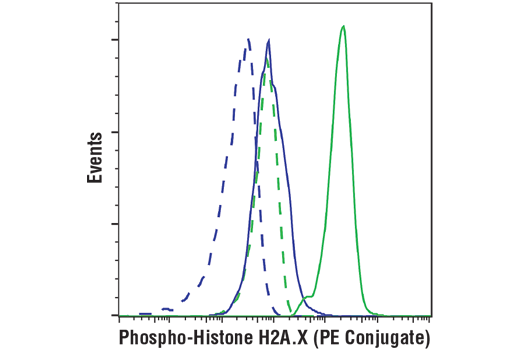
Mouse (G3A1) mAb IgG1 Isotype Control (PE Conjugate) #6899 (dashed lines).
| Cat. # | Size | Qty. | Price |
|---|---|---|---|
| 85410S | 100 µl (50 tests) |
|
| REACTIVITY | H M R Mk |
| SENSITIVITY | Endogenous |
| MW (kDa) | |
| Source/Isotype | Mouse IgG1 |
Product Information
| Application | Dilution |
|---|---|
| Flow Cytometry (Fixed/Permeabilized) | 1:50 |
All reagents required for this protocol may be efficiently purchased together in our Intracellular Flow Cytometry Kit (Methanol) #13593, or individually using the catalog numbers listed below.
NOTE: Prepare solutions with reverse osmosis deionized (RODI) or equivalent grade water.
NOTE: When including fluorescent cellular dyes in your experiment (including viability dyes, DNA dyes, etc.), please refer to the dye product page for the recommended protocol. Visit www.cellsignal.com for a full listing of cellular dyes validated for use in flow cytometry.
NOTE: Adherent cells or tissue should be dissociated and in single-cell suspension prior to fixation.
NOTE: Optimal centrifugation conditions will vary depending upon cell type and reagent volume. Generally, 150-300g for 1-5 minutes will be sufficient to pellet the cells.
NOTE: If using whole blood, lyse red blood cells and wash by centrifugation prior to fixation.
NOTE: Antibodies targeting CD markers or other extracellular proteins may be added prior to fixation if the epitope is disrupted by formaldehyde and/or methanol. The antibodies will remain bound to the target of interest during the fixation and permeabilization process. However, note that some fluorophores (including PE and APC) are damaged by methanol and thus should not be added prior to permeabilization. Conduct a small-scale experiment if you are unsure.
NOTE: Count cells using a hemocytometer or alternative method.
posted July 2009
revised June 2020
Protocol Id: 407
Human, Mouse, Rat, Monkey
Monoclonal antibody is produced by immunizing animals with a synthetic peptide corresponding to residues surrounding Ser139 of human Histone H2A.X protein.
Histone H2A.X is a variant histone that represents approximately 10% of the total H2A histone proteins in normal human fibroblasts (1). H2A.X is required for checkpoint-mediated cell cycle arrest and DNA repair following double-stranded DNA breaks (1). DNA damage, caused by ionizing radiation, UV-light, or radiomimetic agents, results in rapid phosphorylation of H2A.X at Ser139 by PI3K-like kinases, including ATM, ATR, and DNA-PK (2,3). Within minutes following DNA damage, H2A.X is phosphorylated at Ser139 at sites of DNA damage (4). This very early event in the DNA-damage response is required for recruitment of a multitude of DNA-damage response proteins, including MDC1, NBS1, RAD50, MRE11, 53BP1, and BRCA1 (1). In addition to its role in DNA-damage repair, H2A.X is required for DNA fragmentation during apoptosis and is phosphorylated by various kinases in response to apoptotic signals. H2A.X is phosphorylated at Ser139 by DNA-PK in response to cell death receptor activation, c-Jun N-terminal Kinase (JNK1) in response to UV-A irradiation, and p38 MAPK in response to serum starvation (5-8). H2A.X is constitutively phosphorylated on Tyr142 in undamaged cells by WSTF (Williams-Beuren syndrome transcription factor) (9,10). Upon DNA damage, and concurrent with phosphorylation of Ser139, Tyr142 is dephosphorylated at sites of DNA damage by recruited EYA1 and EYA3 phosphatases (9). While phosphorylation at Ser139 facilitates the recruitment of DNA repair proteins and apoptotic proteins to sites of DNA damage, phosphorylation at Tyr142 appears to determine which set of proteins are recruited. Phosphorylation of H2A.X at Tyr142 inhibits the recruitment of DNA repair proteins and promotes binding of pro-apoptotic factors such as JNK1 (9). Mouse embryonic fibroblasts expressing only mutant H2A.X Y142F, which favors recruitment of DNA repair proteins over apoptotic proteins, show a reduced apoptotic response to ionizing radiation (9). Thus, it appears that the balance of H2A.X Tyr142 phosphorylation and dephosphorylation provides a switch mechanism to determine cell fate after DNA damage.
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.