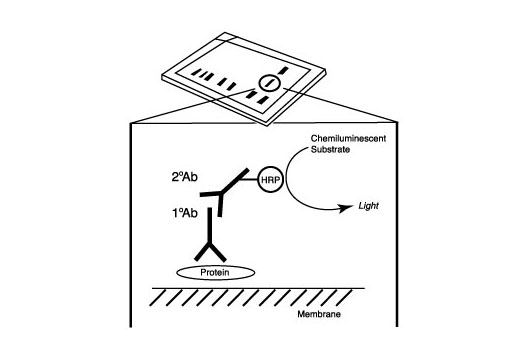
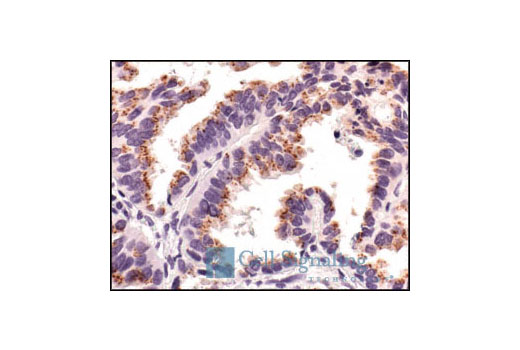
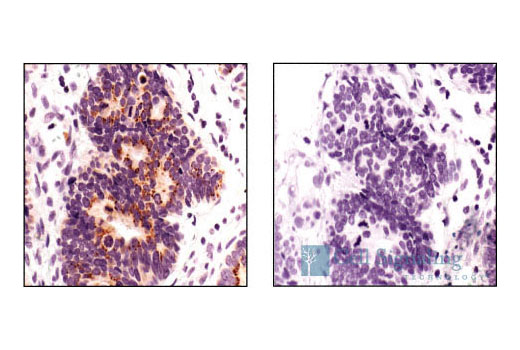
Product Information
Monoclonal antibodies are produced by immunizing animals with a full-length peptide corresponding to human Smac/Diablo protein raised in E. coli, or with synthetic peptides corresponding to amino-terminal residues adjacent to Asp175 of human caspase-3 protein, residues near the carboxy terminus of human COX IV protein, residues surrounding Pro72 of human cytochrome c protein, residues surrounding Phe341 of human HtrA2/Omi protein, or residues surrounding Ala220 of human MEK1 protein.
Apoptosis is a regulated physiological process leading to cell death. Caspases, a family of cysteine acid proteases, are central regulators of apoptosis. Initiator caspases (including 8, 9, 10, and 12) are closely coupled to proapoptotic signals. Once activated, these caspases cleave and activate downstream effector caspases (including 3, 6, and 7), which in turn cleave cytoskeletal and nuclear proteins like PARP, α-fodrin, DFF, and lamin A, and induce apoptosis (1).
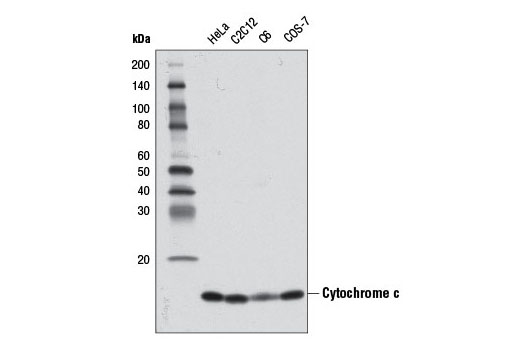
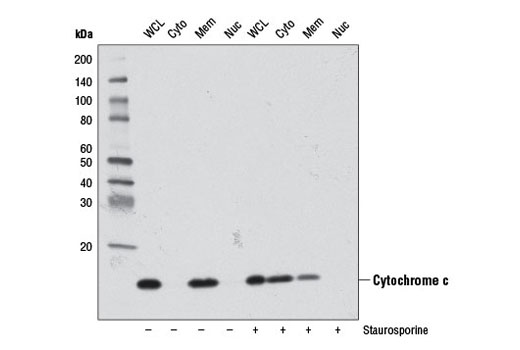
Cytochrome c is a well conserved electron-transport protein and is part of the respiratory chain localized to the mitochondrial intermembrane space (2). Upon apoptotic stimulation, cytochrome c released from mitochondria associates with procaspase-9 (47 kDa)/Apaf-1. This complex processes caspase-9 from inactive proenzyme to its active form (3). This event further triggers caspase-3 activation and eventually leads to apoptosis (4).
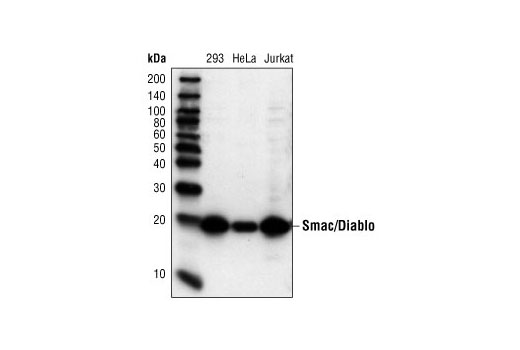
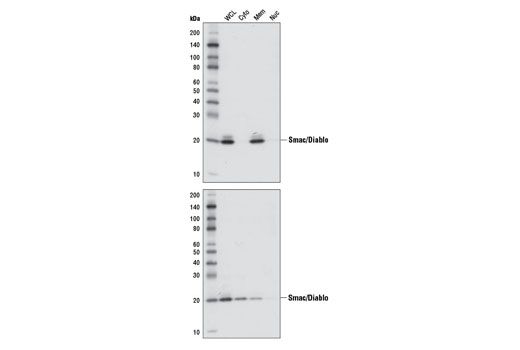
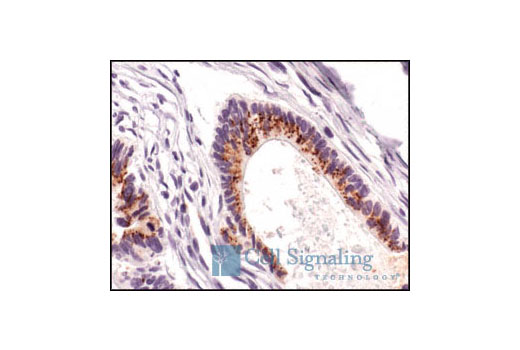
Smac/Diablo is a 21 kDa mammalian mitochondrial protein that functions as a regulatory component during apoptosis (5,6). Upon mitochondrial stress, Smac/Diablo is released from mitochondria and competes with caspases for binding of inhibitor of apoptosis proteins (IAPs) (5,6). The interaction of Smac/Diablo with IAPs relieves the inhibitory effect of the IAPs on caspases (7,8).
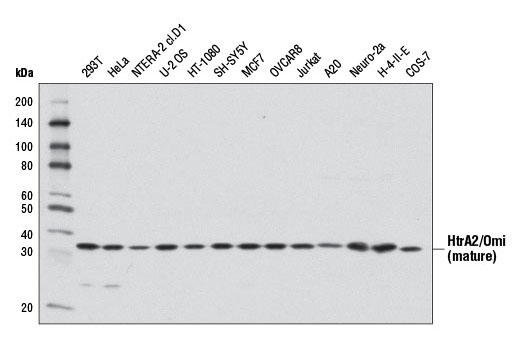
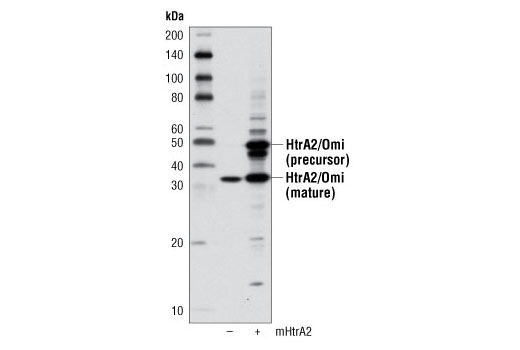
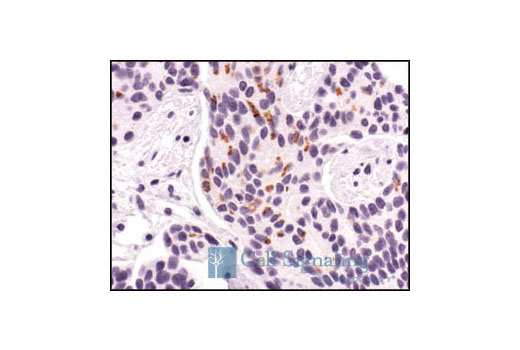
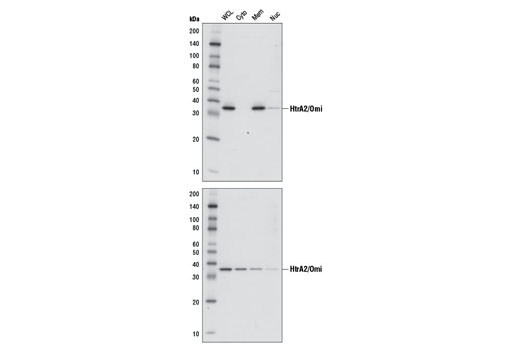
High temperature requirement protein A2 (HtrA2)/Omi is a serine protease with homology to the E. coli HtrA protein (DegP) and is thought to be involved in apoptosis and stress-induced degradation of misfolded proteins (9). HtrA2 is produced as a 50 kDa zymogen that is cleaved to generate a 36 kDa mature protein that exposes an amino terminal motif (AVPS) resembling that of the IAP inhibitor Smac/Diablo (10-14). Like Smac, interaction between HtrA2 and IAP family members, such as XIAP, antagonizes their inhibition of caspase activity and protection from apoptosis (10-14).
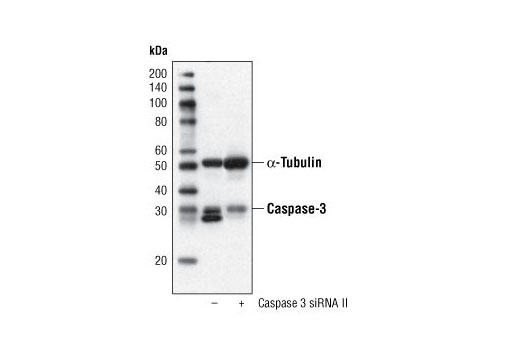
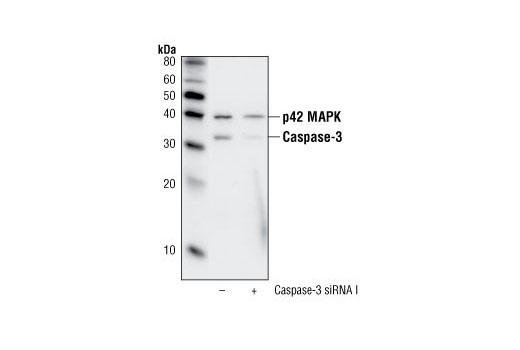
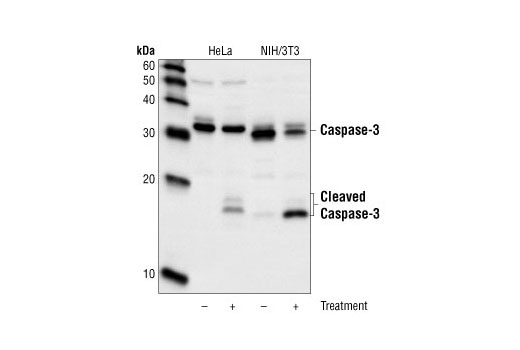
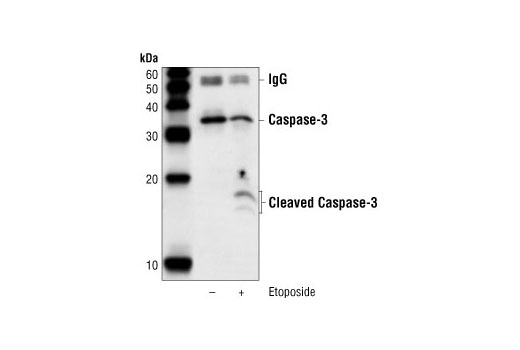
Caspase-3 (CPP-32, Apoptain, Yama, SCA-1) is a critical executioner of apoptosis, as it is either partially or totally responsible for the proteolytic cleavage of many key proteins, such as the nuclear enzyme poly (ADP-ribose) polymerase (PARP) (15). Activation of caspase-3 requires proteolytic processing of its inactive zymogen into activated p17 and p12 fragments. Cleavage of caspase-3 requires the aspartic acid residue at the P1 position (16).
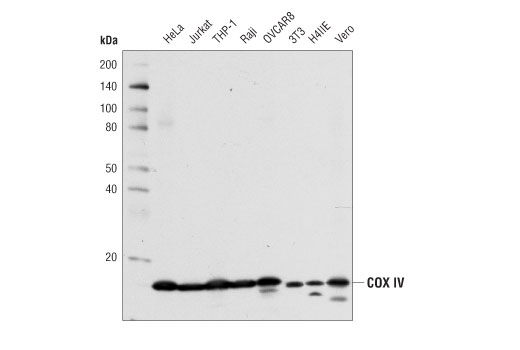
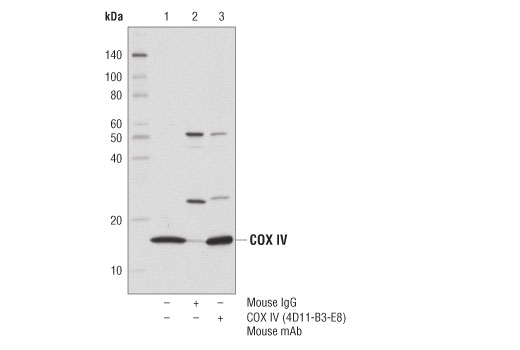
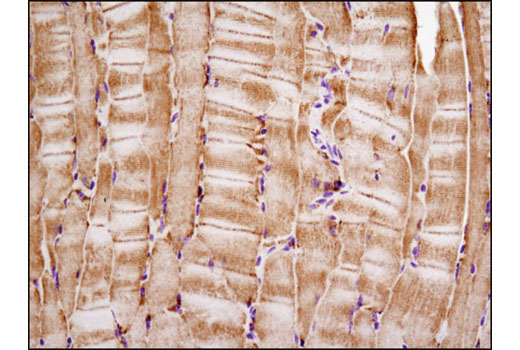
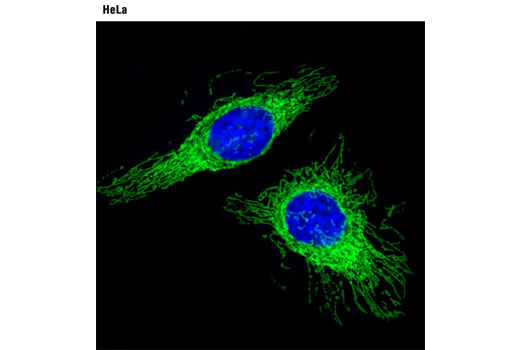
Cytochrome c oxidase (COX) is a hetero-oligomeric enzyme consisting of 13 subunits localized to the inner mitochondrial membrane (17-19). It is the terminal enzyme complex in the respiratory chain, catalyzing the reduction of molecular oxygen to water coupled to the translocation of protons across the mitochondrial inner membrane to drive ATP synthesis. The 3 largest subunits forming the catalytic core are encoded by mitochondrial DNA, while the other smaller subunits, including COX IV, are nuclear-encoded. The COX IV (4D11-B3-E8) Mouse mAb can be used effectively as a mitochondrial loading control in cell-based research assays.
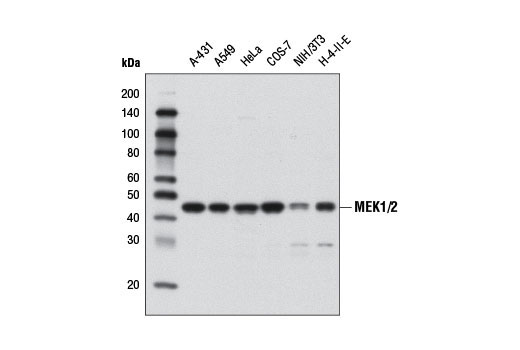
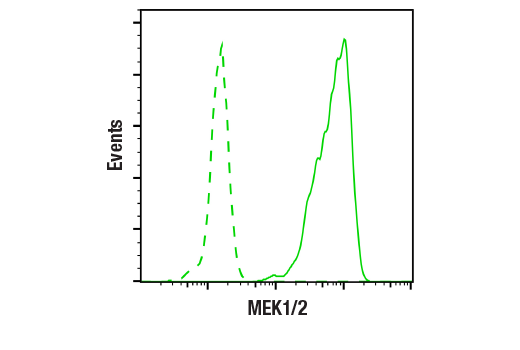
MEK1 and MEK2, also called MAPK or Erk kinases, are dual-specificity protein kinases that function in a mitogen activated protein kinase cascade controlling cell growth and differentiation (20-22). Activation of MEK1 and MEK2 occurs through phosphorylation of two serine residues at positions 217 and 221, located in the activation loop of subdomain VIII, by Raf-like molecules. MEK1/2 is activated by a wide variety of growth factors and cytokines, as well as by membrane depolarization and calcium influx (20-23). MEK activates p44 and p42 MAP kinase by phosphorylating both threonine and tyrosine residues at sites located within the activation loop of kinase subdomain VIII. The MEK1/2 (D1A5) Rabbit mAb can be used effectively as a cytoplasmic loading control in cell-based research assays.
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.