| Cat. # | Size | Qty. | Price |
|---|---|---|---|
| 89985T | 1 Kit (6 x 20 microliters) |
|
| Product Includes | Quantity | Applications | Reactivity | MW(kDa) | Isotype |
|---|---|---|---|---|---|
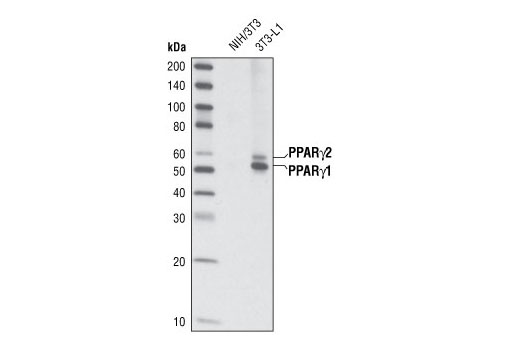
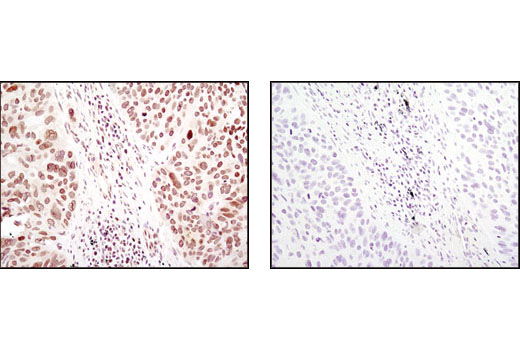
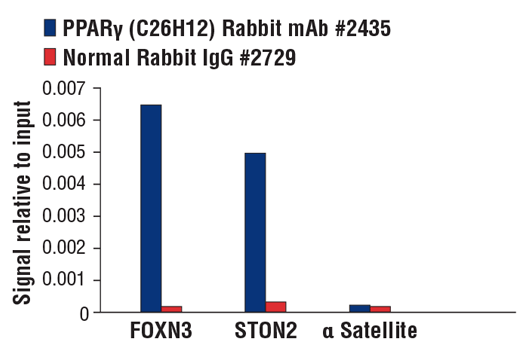
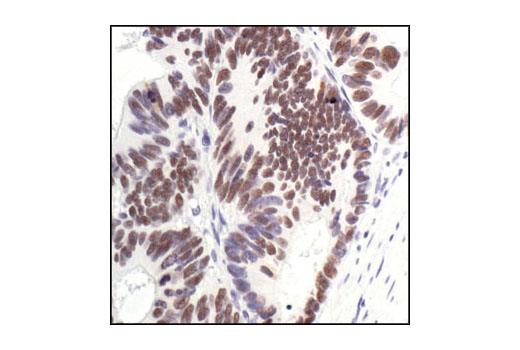
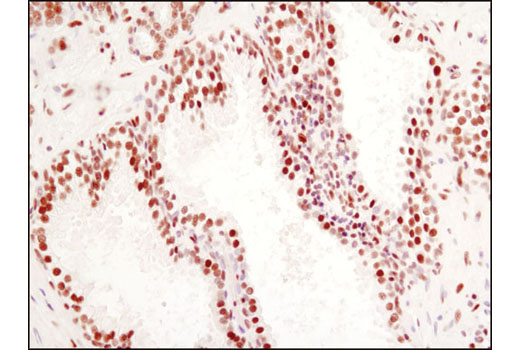
| PPARγ (C26H12) Rabbit mAb 2435 | 20 µl |
|
H M | 53, 57 | Rabbit IgG |
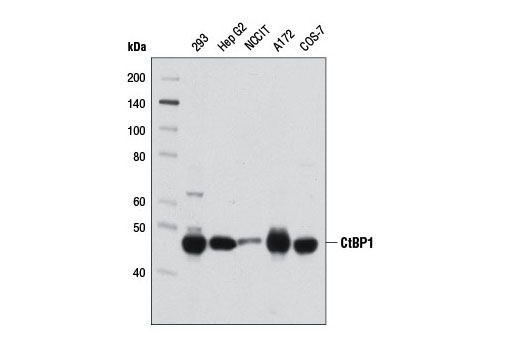
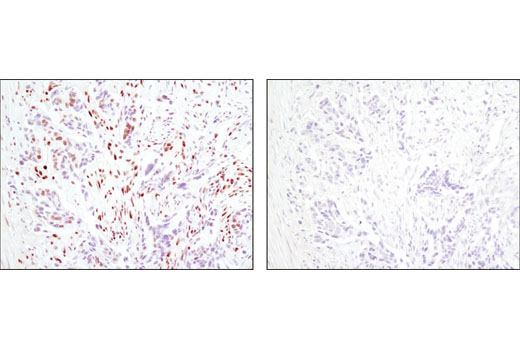
| CtBP1 (D2D6) Rabbit mAb 8684 | 20 µl |
|
H M Mk | 47 | Rabbit IgG |
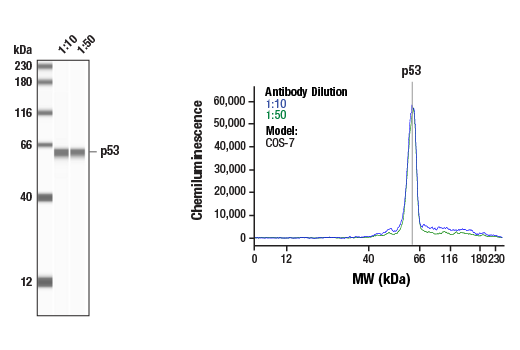
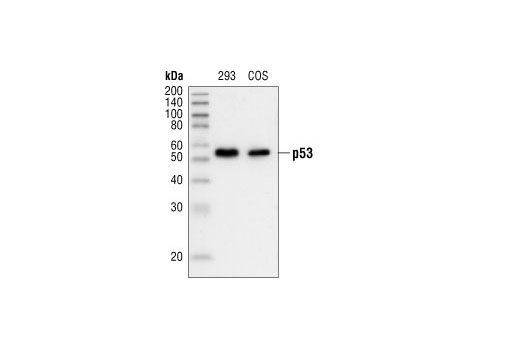
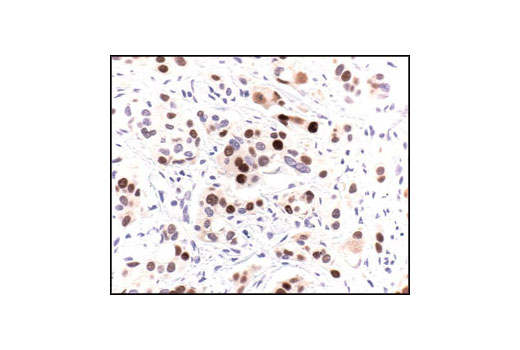
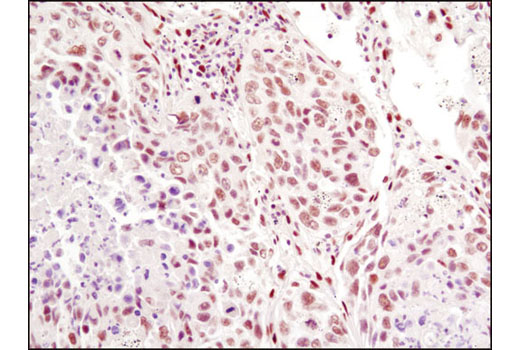
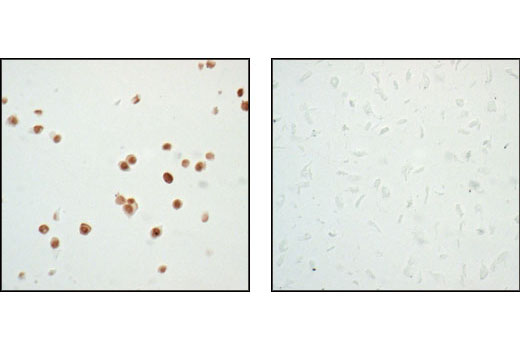
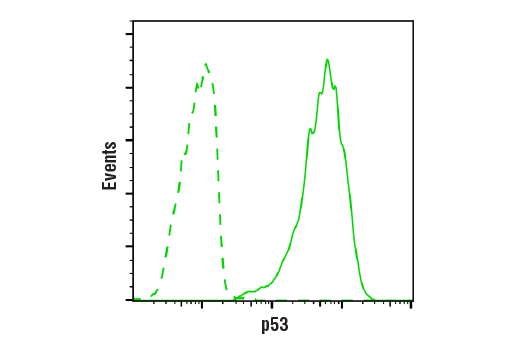
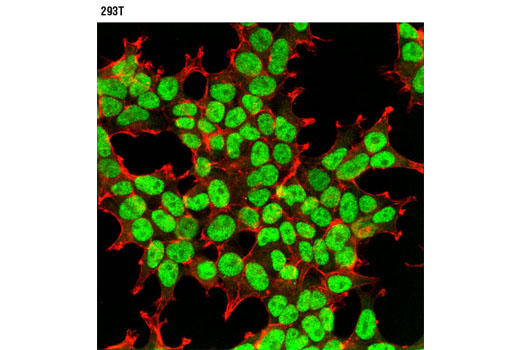
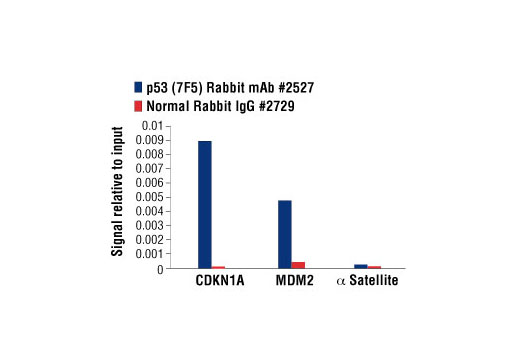
| p53 (7F5) Rabbit mAb 2527 | 20 µl |
|
H Mk | 53 | Rabbit IgG |
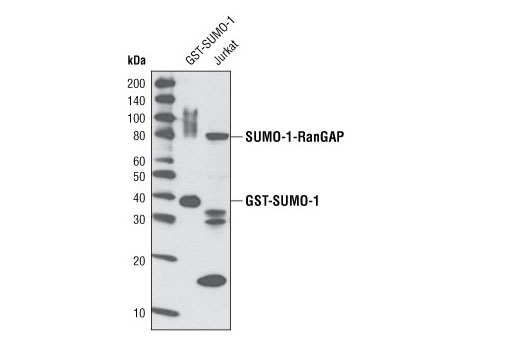
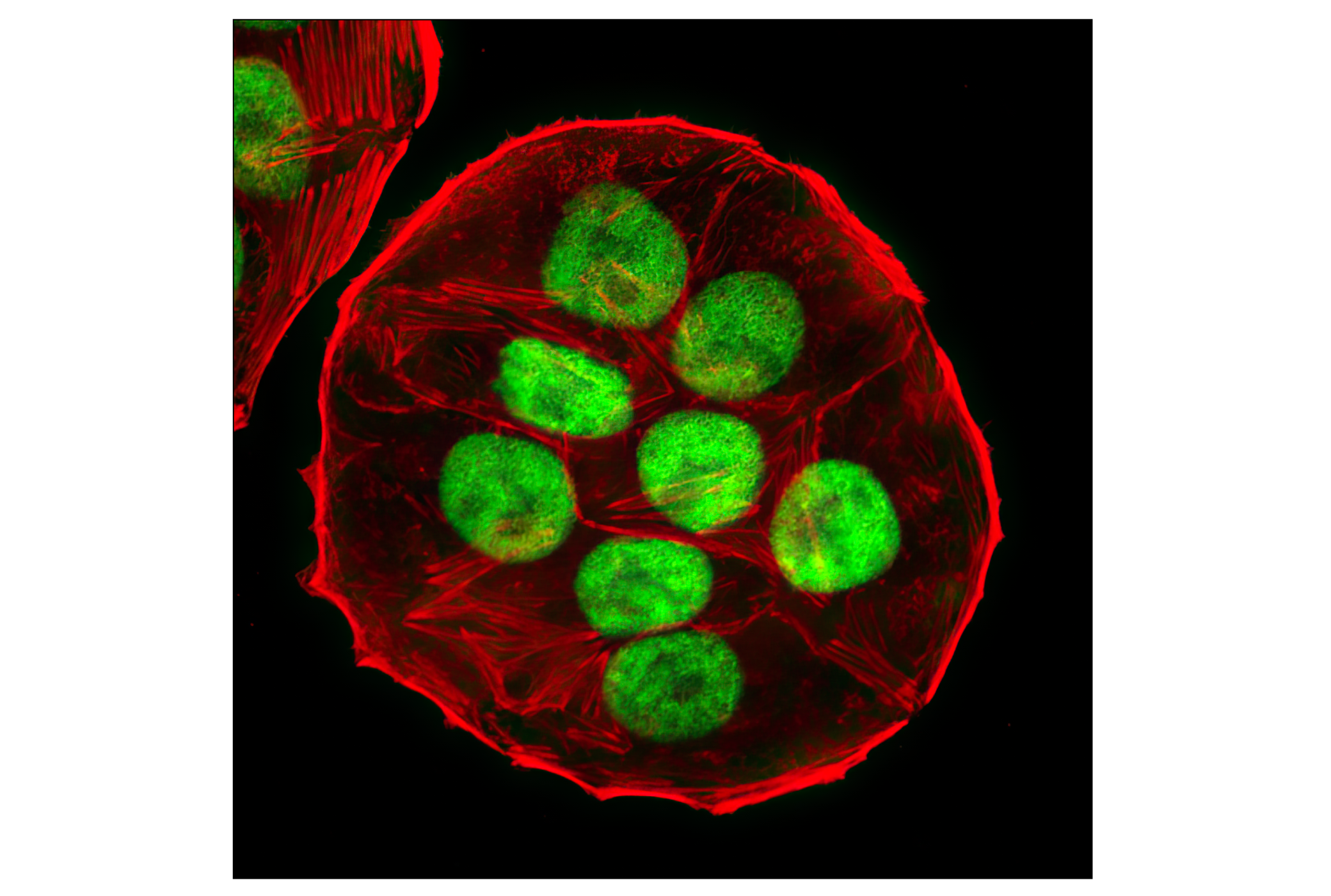
| SUMO-1 (C9H1) Rabbit mAb 4940 | 20 µl |
|
H M R Mk | Rabbit IgG | |
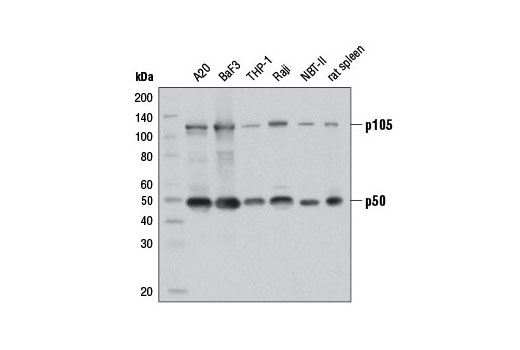
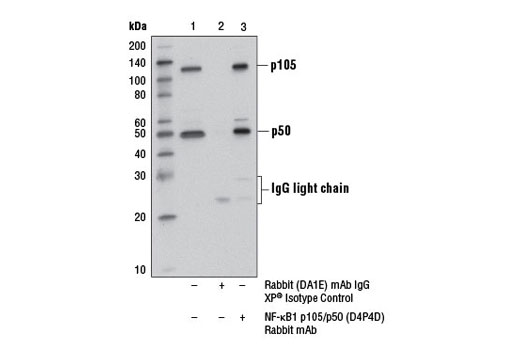
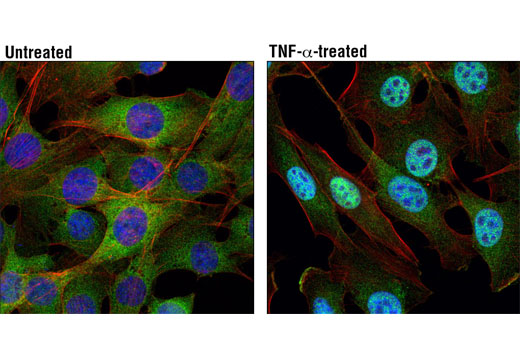
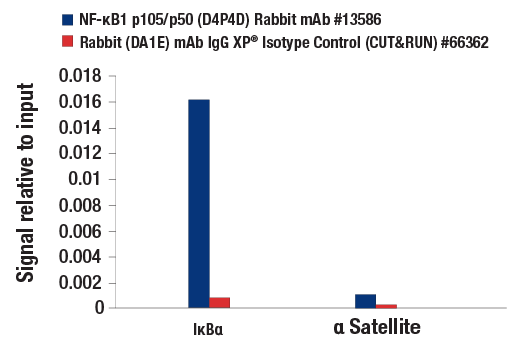
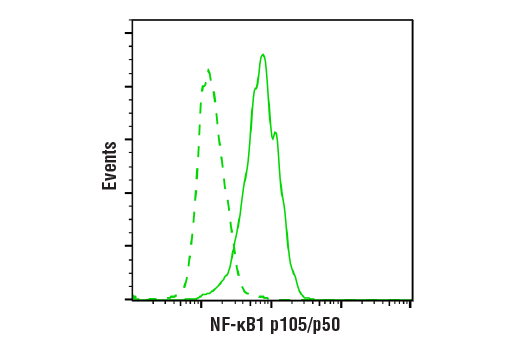
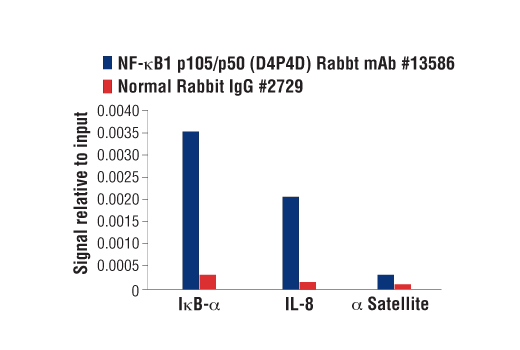
| NF-κB1 p105/p50 (D4P4D) Rabbit mAb 13586 | 20 µl |
|
H M R | 50 Active form. 120 Precursor | Rabbit IgG |
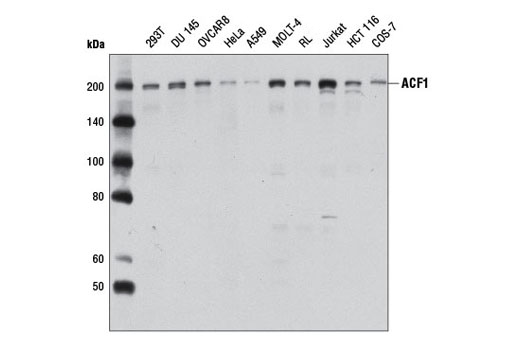
| ACF1 Antibody 6255 | 20 µl |
|
H Mk | 203 | Rabbit |
| Anti-rabbit IgG, HRP-linked Antibody 7074 | 100 µl |
|
Rab | Goat |
Product Information
Monoclonal antibodies are produced by immunizing animals with a synthetic peptide corresponding to the residues surrounding Asp69 of human PPARγ, amino terminus of the human CtBP1 protein, full-length human p53 fusion protein, amino terminus of human SUMO-1, or residues surrounding Ile415 of mouse NF-kB1 P105/p50 protein. Polyclonal antibodies are produced by immunizing animals with a synthetic peptide corresponds to the residues surrounding Met864 of human ACF1 protein. Antibodies are purified by protein A and peptide affinity chromatography
Peroxisome proliferator-activated receptor gamma (PPARG) is a member of the ligand-activated nuclear receptor superfamily and functions as a transcriptional activator (1). Besides its role in mediating adipogenesis and lipid metabolism (2), PPAR gamma also modulates insulin sensitivity, cell proliferation and inflammation (3). CtBP1 is able to regulate gene activity through its intrinsic dehydrogenase activity (4,5) and by interacting with Polycomb Group (PcG) proteins during development (6). Along with its homologue, CtBP2, it acts as a transcriptional corepressor of zinc-finger homeodomain factor deltaEF1 to regulate a wide range of cellular processes through transrepression mechanisms (7). The p53 tumor suppressor protein plays a major role in cellular response to DNA damage and other genomic aberrations. Activation of p53 can lead to either cell cycle arrest and DNA repair or apoptosis (8). DNA damage induces phosphorylation of p53 at Ser15 and Ser20 and leads to a reduced interaction between p53 and its negative regulator, the oncoprotein MDM2 (9). MDM2 inhibits p53 accumulation by targeting it for ubiquitination and proteasomal degradation (10,11). Phosphorylation impairs the ability of MDM2 to bind p53, promoting both the accumulation and activation of p53 in response to DNA damage (9,12). Acetylation appears to play a positive role in the accumulation of p53 protein in stress response (13). Deacetylation of p53 occurs through interaction with the SIRT1 protein, a deacetylase that may be involved in cellular aging and the DNA damage response (14). Small ubiquitin-related modifier 1, 2 and 3 (SUMO-1, -2 and -3) are members of the ubiquitin-like protein family (15). The covalent attachment of the SUMO-1, -2 or -3 (SUMOylation) to target proteins is analogous to ubiquitination. Ubiquitin and the individual SUMO family members are all targeted to different proteins with diverse biological functions. Ubiquitin predominantly regulates degradation of its target (1). In contrast, SUMO-1 is conjugated to RanGAP, PML, p53 and IkB-alpha to regulate nuclear trafficking, formation of subnuclear structures, regulation of transcriptional activity and protein stability (16-20). Transcription factors of the nuclear factor kappaB (NF-kB)/Rel family play a pivotal role in inflammatory and immune responses (21, 22). In unstimulated cells, NF-kB is sequestered in the cytoplasm by IkB inhibitory proteins (23-25). NF-kB-activating agents can induce the phosphorylation of IkB proteins, targeting them for rapid degradation through the ubiquitin-proteasome pathway and releasing NF-kB to enter the nucleus where it regulates gene expression (26-28). ACF1 (BAZ1A) has distinct roles in development (29), regulation of chromatin structure (30), and DNA damage response (31, 32). Different developmental stages dictate the expression of ACF1 in Drosophila, and alterations in ACF1 expression during Drosophila development leads to deviation from normal chromatin organization (29).
Explore pathways related to this product.
STRING - Known and Predicted Protein-Protein Interactions.
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.