| Cat. # | Size | Qty. | Price |
|---|---|---|---|
| 8658T | 1 Kit (9 x 20 microliters) |
|
| Product Includes | Quantity | Applications | Reactivity | MW(kDa) | Isotype |
|---|---|---|---|---|---|
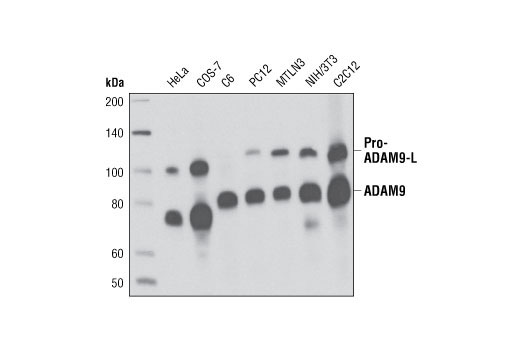
| ADAM9 (D64B5) Rabbit mAb 4151 | 20 µl |
|
H M R Mk | 100-115, 75-80 | Rabbit IgG |
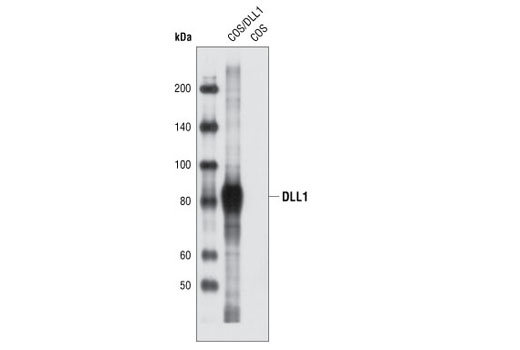
| DLL1 Antibody 2588 | 20 µl |
|
R | 82 | Rabbit |
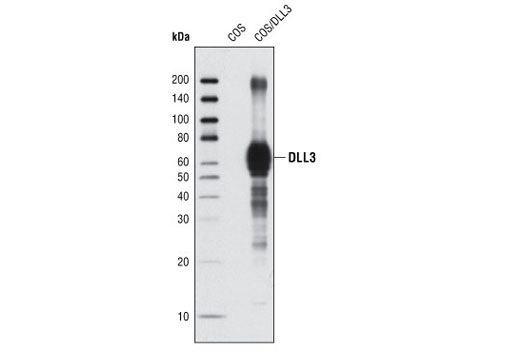
| DLL3 (G93) Antibody 2483 | 20 µl |
|
R | 65 | Rabbit |
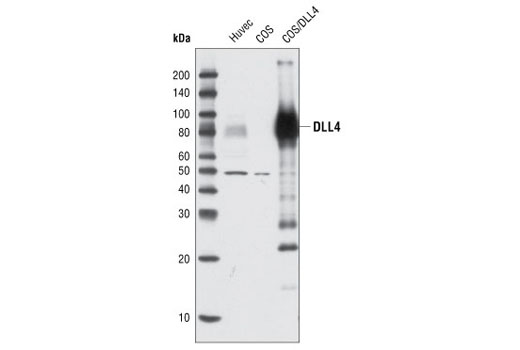
| DLL4 Antibody 2589 | 20 µl |
|
H | 75-80 | Rabbit |
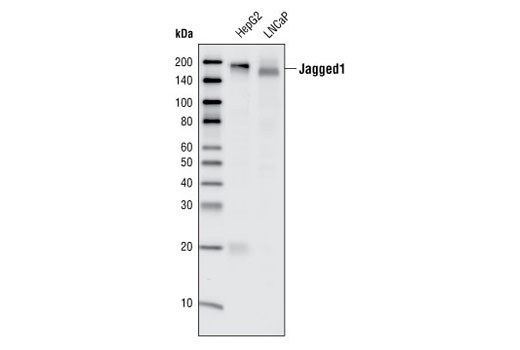
| Jagged1 (28H8) Rabbit mAb 2620 | 20 µl |
|
H M | 180 | Rabbit IgG |
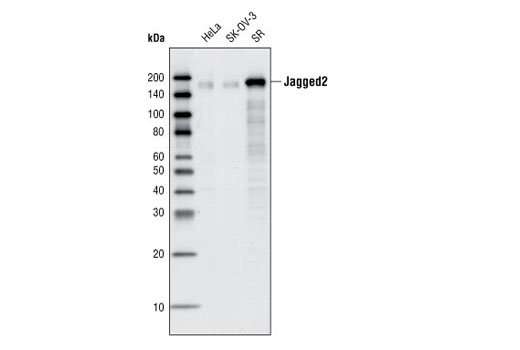
| Jagged2 (C23D2) Rabbit mAb 2210 | 20 µl |
|
H R | 150 | Rabbit IgG |
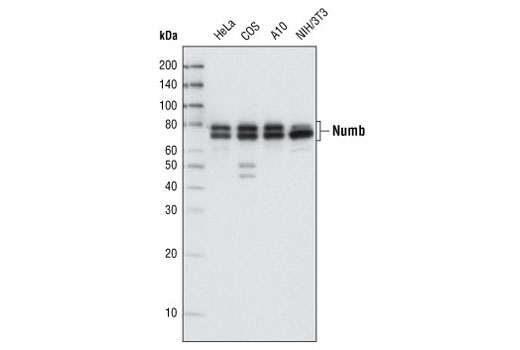
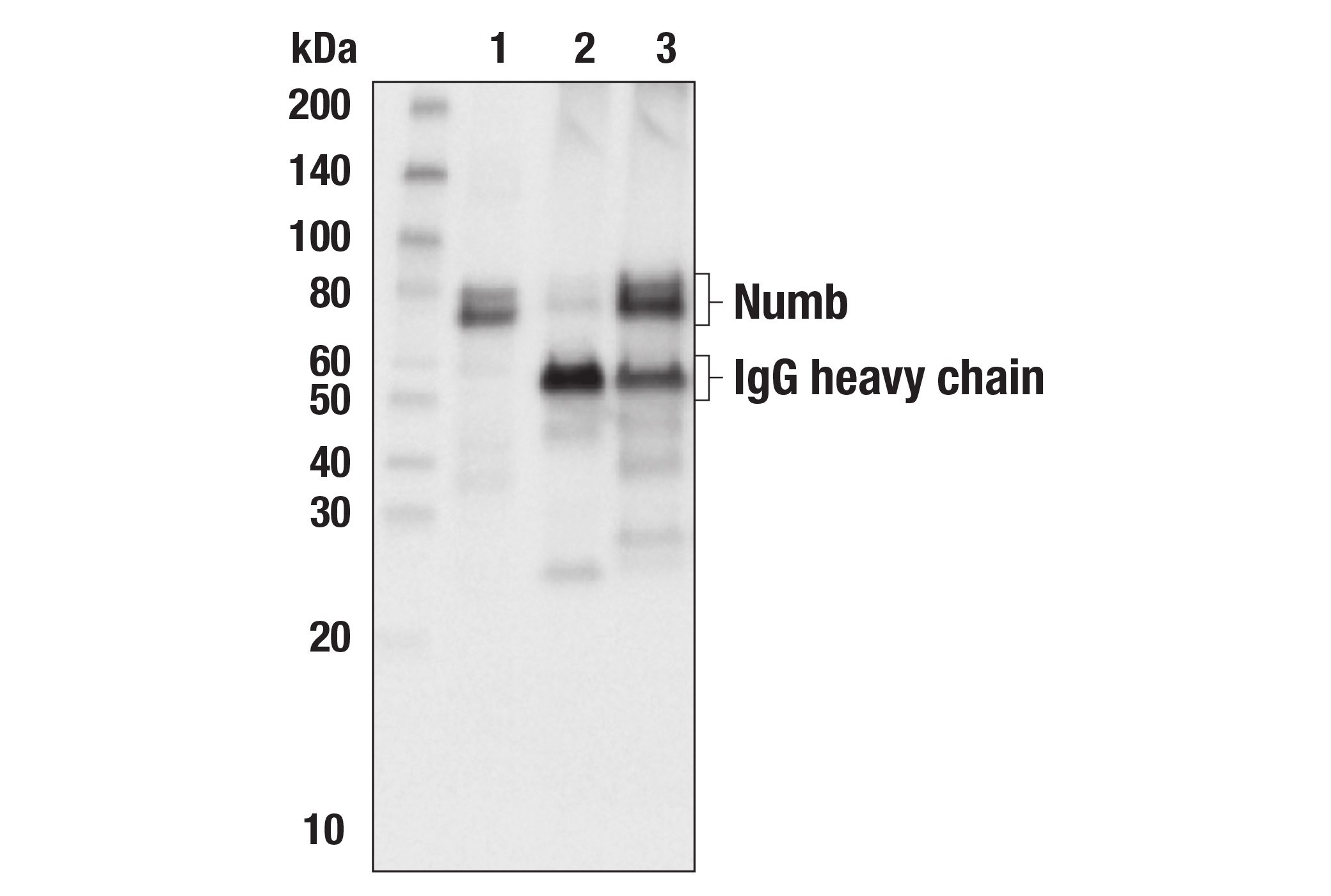
| Numb (C29G11) Rabbit mAb 2756 | 20 µl |
|
H M R Mk | 72, 74 | Rabbit IgG |
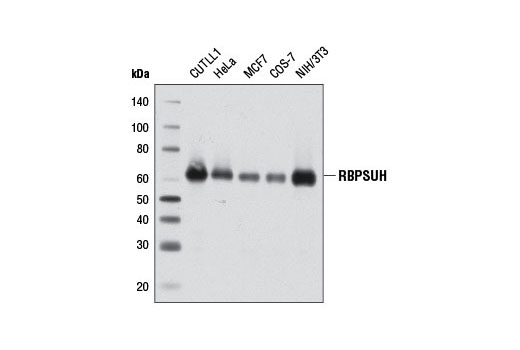
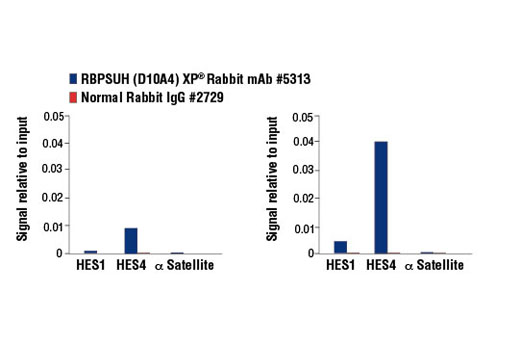
| RBPSUH (D10A4) XP® Rabbit mAb 5313 | 20 µl |
|
H M R Mk | 61 | Rabbit IgG |
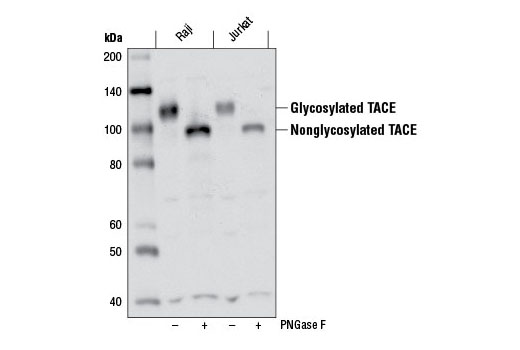
| TACE (D22H4) Rabbit mAb 6978 | 20 µl |
|
H | 135 | Rabbit IgG |
| Anti-rabbit IgG, HRP-linked Antibody 7074 | 100 µl |
|
Rab | Goat |
Product Information
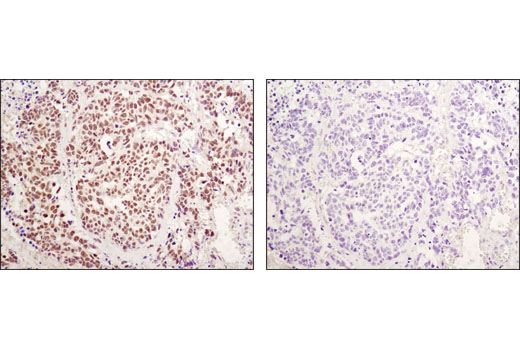
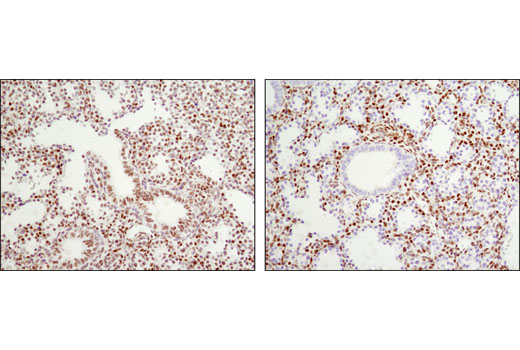
Monoclonal antibodies are produced by immunizing animals either with a recombinant protein specific to the amino terminus of human TACE protein or with a synthetic peptide corresponding to residues surrounding Glu1140 (intracellular region) of human Jagged1 protein, residues surrounding Ala117 of human Jagged2 protein, residues surrounding Ala570 of human Numb protein, or residues near the carboxy terminus of human ADAM9 protein or residues surrounding Gln110 of human RBPSUH protein.
Polyclonal antibodies are produced by immunizing animals with a synthetic peptide corresponding to residues surrounding Ala627 of human DLL1 protein, residues surrounding Gly93 of mouse DLL3 protein, residues surrounding Leu617 of human DLL4 protein. Polyclonal antibodies are purified by protein A and peptide affinity chromatography.
Notch signaling is activated upon engagement of the Notch receptor with its ligands, the Delta, Serrate, Lag2 (DSL) single-pass type I membrane proteins. DSL proteins contain multiple EGF-like repeats and a DSL domain that is required for binding to Notch (1,2). Five DSL proteins have been identified in mammals: Jagged1, Jagged2, Delta-like (DLL) 1, 3, and 4 (3). Ligand binding to the Notch receptor results in two sequential proteolytic cleavages of the receptor by the ADAM protease and the γ-secretase complex. The intracellular domain of Notch is released and then translocates to the nucleus where it activates transcription. Notch ligands may also be processed in a similiar manner, suggesting bi-directional signaling through receptor-ligand interactions (4-6).
TNF-α converting enzyme (TACE), also known as ADAM17, is a transmembrane metalloprotease that plays a key role in the cleavage of a number cell surface molecules in a process known as “shedding". TACE is abundantly expressed in many adult tissues, but in fetal development, expression is differentially regulated (7). TACE activates Notch in a ligand-independent manner and has been shown to play a role in the development of the Drosophila nervous system (8).
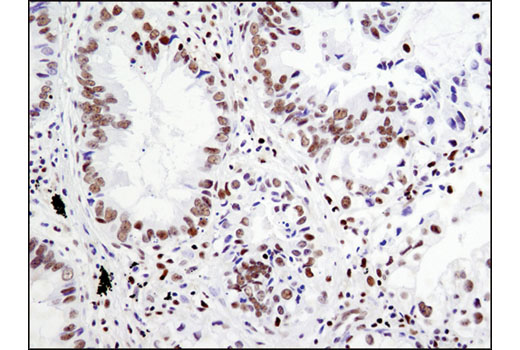
Recombining Binding Protein, SUppressor of Hairless (RBPSUH), also termed RBP-J or CSL, is the DNA-binding component of the transcription complex regulated by canonical Notch signaling. In the absence of Notch activation, RBPSUH suppresses target gene expression through interactions with a co-repressor complex containing histone deacetylase. Upon activation of Notch receptors, the Notch intracellular domain (NICD) translocates to the nucleus and binds to RBPSUH. This displaces the co-repressor complex and replaces it with a transcription activation complex that includes Mastermind-like (MAML) proteins and histone acetylase p300, leading to transcriptional activation of Notch target genes (9-11).
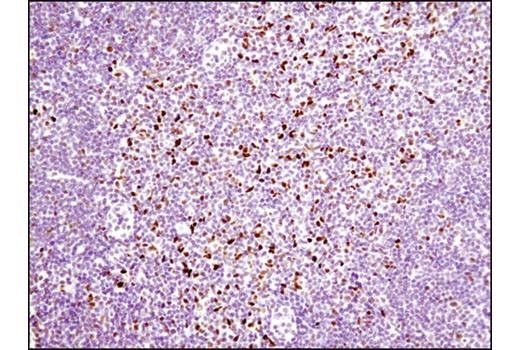
Numb contains an amino-terminal phosphotyrosine-binding (PTB) domain and carboxy-terminal endocytic binding motifs for α-adaptin and EH (Eps15 homology) domain-containing proteins, indicating a role in endocytosis (12,13). There are four mammalian Numb splicing isoforms that are differentially expressed and may have distinct functions (14-16). Numb acts as a negative regulator of Notch signaling by promoting ubiquitination and degradation of Notch (17). The protein is asymmetrically segregated into one daughter cell during cell division, producing two daughter cells with different responses to Notch signaling and different cell fates (18,19).
Explore pathways related to this product.
STRING - Known and Predicted Protein-Protein Interactions.
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.