| REACTIVITY | H |
Product Information
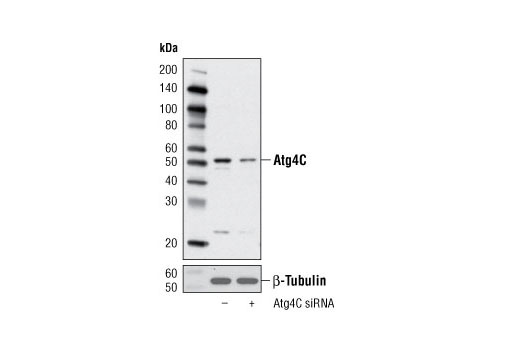
CST recommends transfection with 100 nM Atg4C siRNA I 48 to 72 hours prior to cell lysis. For transfection procedure, follow protocol provided by the transfection reagent manufacturer. Please feel free to contact CST with any questions on use.
Each vial contains the equivalent of 100 transfections, which corresponds to a final siRNA concentration of 100 nM per transfection in a 24-well plate with a total volume of 300 μl per well.
Oligonucleotide synthesis is monitored base by base through trityl analysis to ensure appropriate coupling efficiency. The oligo is subsequently purified by affinity-solid phase extraction. The annealed RNA duplex is further analyzed by mass spectrometry to verify the exact composition of the duplex. Each lot is compared to the previous lot by mass spectrometry to ensure maximum lot-to-lot consistency.
Autophagy is a catabolic process for the autophagosomic-lysosomal degradation of bulk cytoplasmic contents. Control of autophagy was largely discovered in yeast and involves proteins encoded by a set of autophagy-related genes (Atg) (1). Formation of autophagic vesicles requires a pair of essential ubiquitin-like conjugation systems, Atg12-Atg5 and Atg8-phosphatidylethanolamine (Atg8-PE), which are widely conserved in eukaryotes (2). Numerous mammalian counterparts to yeast Atg proteins have been described, including three Atg8 proteins (GATE-16, GABARAP, and LC3) and four Atg4 homologs (Atg4A/autophagin-2, Atg4B/autophagin-1, Atg4C/autophagin-3, and Atg4D/autophagin-4) (3-5). The cysteine protease Atg4 is pivotal to autophagosome membrane generation and regulation. Atg4 primes the Atg8 homolog for lipidation by cleaving its carboxy terminus and exposing its glycine residue for E1-like enzyme Atg7. The Atg8 homolog is transferred to the E2-like enzyme Atg3 before forming the Atg8-PE conjugate. During later stages of autophagy, Atg4 can reverse this lipidation event by cleaving PE, thereby recycling the Atg8 homolog (6).
Atg4C-deficient mice display a tissue-specific decrease in LC3 lipidation only when under stressful conditions such as prolonged starvation. Mutant mice also exhibit increased susceptibility to the development of chemical carcinogen induced fibrosarcomas suggesting that Atg4C may contribute to events associated with tumor progression (7).
Explore pathways related to this product.
STRING - Known and Predicted Protein-Protein Interactions.
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.