| REACTIVITY | H |
Product Information
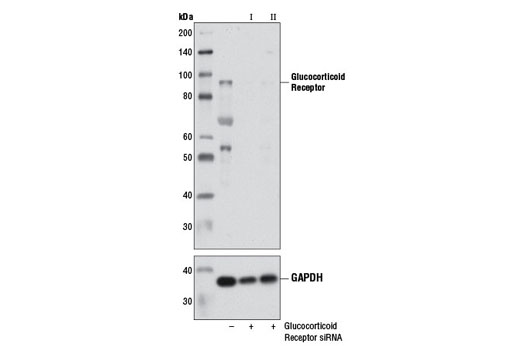
CST recommends transfection with 100 nM SignalSilence® Glucocorticoid Recpetor siRNA I 48 to 72 hours prior to cell lysis. For transfection procedure, follow protocol provided by the transfection reagent manufacturer. Please feel free to contact CST with any questions on use.
Each vial contains the equivalent of 100 transfections, which corresponds to a final siRNA concentration of 100 nM per transfection in a 24-well plate with a total volume of 300 μl per well.
Oligonucleotide synthesis is monitored base by base through trityl analysis to ensure appropriate coupling efficiency. The oligo is subsequently purified by affinity-solid phase extraction. The annealed RNA duplex is further analyzed by mass spectrometry to verify the exact composition of the duplex. Each lot is compared to the previous lot by mass spectrometry to ensure maximum lot-to-lot consistency.
Glucocorticoid hormones control cellular proliferation, inflammation, and metabolism through their association with the glucocorticoid receptor (GR)/NR3C1, a member of the nuclear hormone receptor superfamily of transcription factors (1). GR is composed of several conserved structural elements, including a carboxy-terminal ligand-binding domain (which also contains residues critical for receptor dimerization and hormone-dependent gene transactivation), a neighboring hinge region containing nuclear localization signals, a central zinc-finger-containing DNA-binding domain, and an amino-terminal variable region that participates in ligand-independent gene transcription. In the absence of hormone, a significant population of GR is localized to the cytoplasm in an inactive form via its association with regulatory chaperone proteins, such as HSP90, HSP70, and FKBP52. On hormone binding, GR is released from the chaperone complex and translocates to the nucleus as a dimer to associate with specific DNA sequences termed glucocorticoid response elements (GREs), thereby enhancing or repressing transcription of specific target genes (2). It was demonstrated that GR-mediated transcriptional activation is modulated by phosphorylation (3-5). Although GR can be basally phosphorylated in the absence of hormone, it becomes hyperphosphorylated upon binding receptor agonists. It has been suggested that hormone-dependent phosphorylation of GR may determine target promoter specificity, cofactor interaction, strength and duration of receptor signaling, receptor stability, and receptor subcellular localization (3).
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.
| Cat. # | Size | Qty. | Price |
|---|---|---|---|
| 6508S | 300 µl (3 nmol) |
|