

Epigenetics
CST signaling pathway diagrams allow you to click on individual nodes to find research resources or product information. You can also download the pathway diagrams for educational and research purposes.
An Introduction to Epigenetics Video
Chromatin regulation refers to the events affecting chromatin structure and therefore, transcriptional control of gene expression patterns.
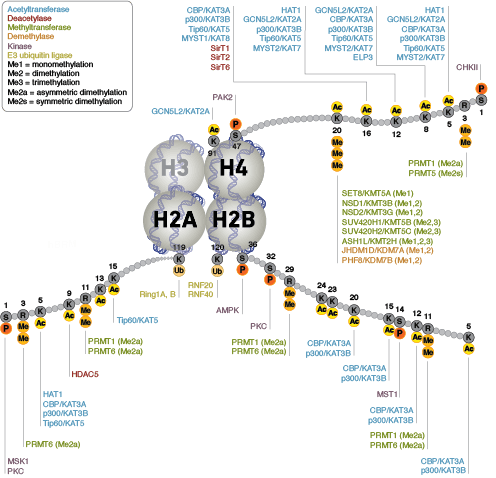
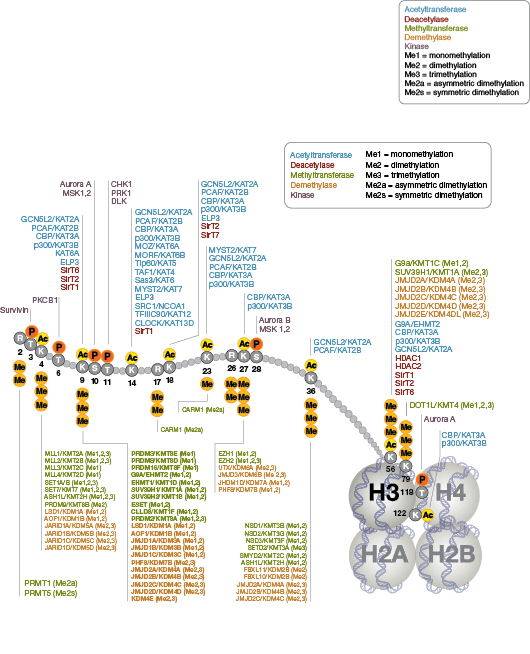
Epigenetics, specifically, refers to the heritable modifications which result in altered gene expression and are not known to be encoded in DNA. The nucleosome, made up of four histone proteins (H2A, H2B, H3, and H4), is the primary building block of chromatin. Originally thought to function as a static scaffold for DNA packaging, histones have more recently been shown to be dynamic proteins, undergoing multiple types of post-translational modifications (PTMs) and interacting with regulatory proteins to control gene expression.
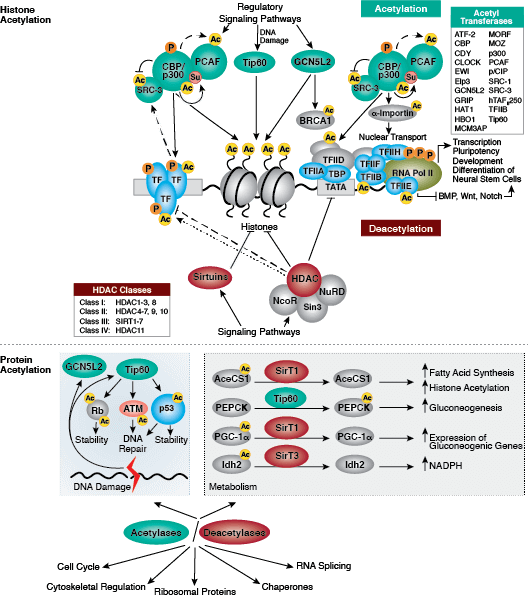
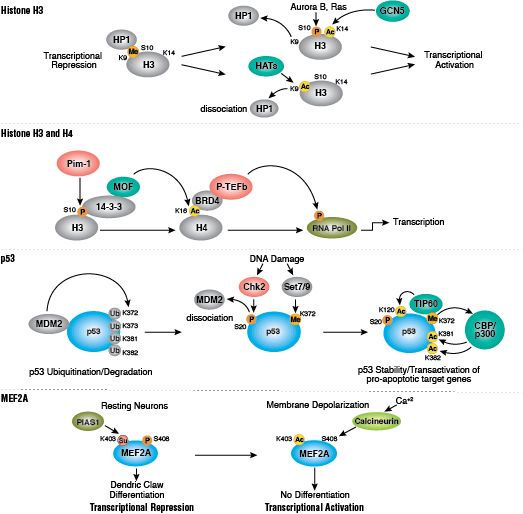
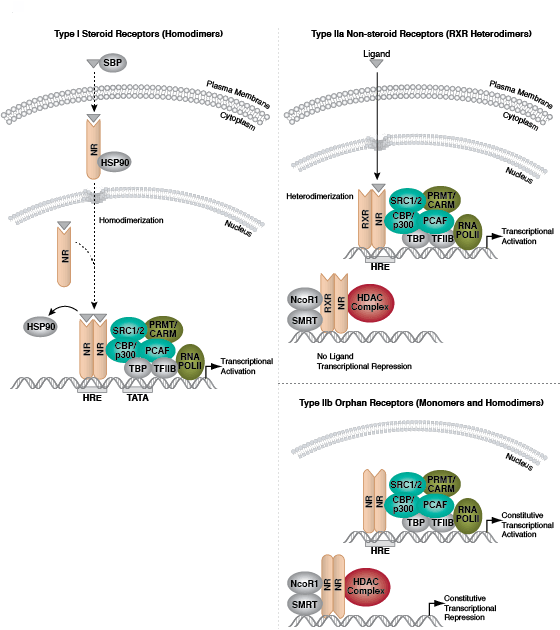
Protein acetylation plays a crucial role in regulating chromatin structure and transcriptional activity. Histone hyperacetylation by histone acetyltransferases (HATs) is associated with transcriptional activation, whereas histone deacetylation by histone deacetylases (HDACs) is associated with transcriptional repression. Hyperacetylation can directly affect chromatin structure by neutralizing the positive charge on histone tails and disrupting nucleosome-nucleosome and nucleosome-DNA interactions. In addition, acetylation creates binding sites for bromodomain-containing chromatin regulatory proteins (histone modification readers).
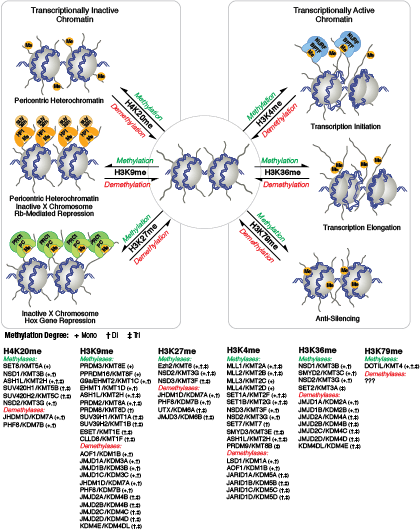
Unlike acetylation, methylation does not alter the charge of arginine and lysine residues and is unlikely to directly modulate nucleosomal interactions required for chromatin folding. Methylated arginine and lysine residues are major determinants for formation of active and inactive regions of the genome. Methylation facilitates binding of chromatin regulatory proteins/histone modification readers that contain various methyl-lysine or methyl-arginine binding domains (PHD, chromo, WD40, Tudor, MBT, Ankyrin repeats, PWWP domains). Recruitment of co-activator and co-repressor proteins is dependent on the specific lysine residue that is modified.
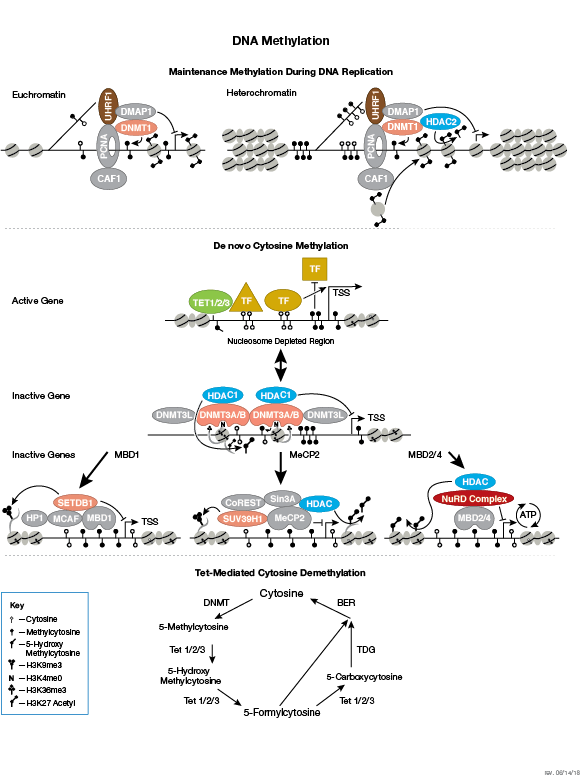
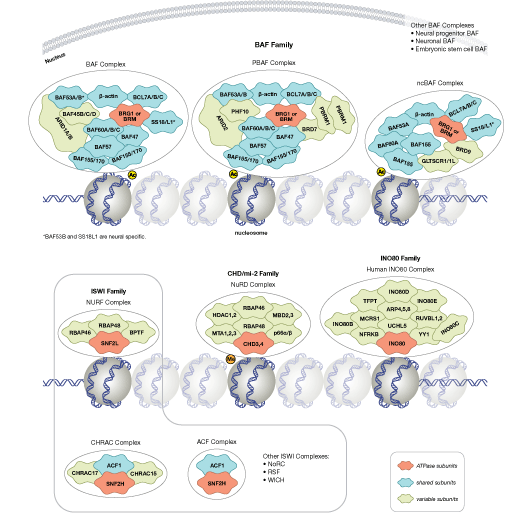
The modulation of chromatin structure is an essential component in the regulation of transcriptional activation and repression. One strategy by which chromatin structure can be modulated is through disruption of histone-DNA contacts by ATP-dependent chromatin remodelers, such as the NuRD, Polycomb, and SWI/SNF complexes, which have been shown to regulate gene activation/repression, cell growth, the cell cycle, and differentiation. Chromatin structure is also modulated through other PTMs such as phosphorylation of histone proteins, which affects association with DNA-interacting proteins and has been recently identified to play a role in coordinating other histone modifications. Furthermore, methylation of DNA at cytosine residues in mammalian cells affects chromatin folding and is a heritable, epigenetic modification that is critical for proper regulation of gene silencing, genomic imprinting, and development. Three families of mammalian DNA methyl-transferases have been identified, DNMT1/2/3, that play distinct roles in embryonic stem cells and adult somatic cells. In addition to the core histone proteins, a number of histone variants exist that confer different structural properties to nucleosomes and play a number of specific functions such as DNA repair, proper kinetochore assembly and chromosome segregation during mitosis, and regulation of transcription.
Chromatin and epigenetic regulation is crucial for proper programming of the genome during development and under stress conditions, as the misregulation of gene expression can lead to diseased states such as cancer.
References:
- Li X, Zhao X (2008) Epigenetic regulation of mammalian stem cells. Stem Cells Dev. 17(6), 1043–52.
- Shilatifard A (2008) Molecular implementation and physiological roles for histone H3 lysine 4 (H3K4) methylation. Curr. Opin. Cell Biol. 20(3), 341–8.
- Haberland M, Montgomery RL, Olson EN (2009) The many roles of histone deacetylases in development and physiology: implications for disease and therapy. Nat. Rev. Genet. 10(1), 32–42.
- Wu SC, Zhang Y (2010) Active DNA demethylation: many roads lead to Rome. Nat. Rev. Mol. Cell Biol. 11(9), 607–20.
- Banaszynski LA, Allis CD, Lewis PW (2010) Histone variants in metazoan development. Dev. Cell 19(5), 662–74.
- Yun M, Wu J, Workman JL, Li B (2011) Readers of histone modifications. Cell Res. 21(4), 564–78.
- Chen ZX, Riggs AD (2011) DNA methylation and demethylation in mammals. J. Biol. Chem. 286(21), 18347–53.
- Lai AY, Wade PA (2011) Cancer biology and NuRD: a multifaceted chromatin remodelling complex. Nat. Rev. Cancer 11(8), 588–96.
- Magnani L, Eeckhoute J, Lupien M (2011) Pioneer factors: directing transcriptional regulators within the chromatin environment. Trends Genet. 27(11), 465–74.
- Zhou Y, Kim J, Yuan X, Braun T (2011) Epigenetic modifications of stem cells: a paradigm for the control of cardiac progenitor cells. Circ. Res. 109(9), 1067–81.
- Banerjee T, Chakravarti D (2011) A peek into the complex realm of histone phosphorylation. Mol. Cell. Biol. 31(24), 4858–73.
- de Nadal E, Ammerer G, Posas F (2011) Controlling gene expression in response to stress. Nat. Rev. Genet. 12(12), 833–45.