| # | Product Name | Applications | Reactivity | |
|---|---|---|---|---|
| 27683 | Acetyl-Histone H3 (Lys36) (D9T5Q) Rabbit mAb |
|
H M R Mk |
| REACTIVITY | H M R Mk |
| SENSITIVITY | Endogenous |
| MW (kDa) | 17 |
| SOURCE | Rabbit |
Product Information
| Application | Dilution |
|---|---|
| Western Blotting | 1:1000 |
For western blots, incubate membrane with diluted primary antibody in 5% w/v BSA, 1X TBS, 0.1% Tween® 20 at 4°C with gentle shaking, overnight.
NOTE: Please refer to primary antibody product webpage for recommended antibody dilution.
From sample preparation to detection, the reagents you need for your Western Blot are now in one convenient kit: #12957 Western Blotting Application Solutions Kit
NOTE: Prepare solutions with reverse osmosis deionized (RODI) or equivalent grade water.
Load 20 µl onto SDS-PAGE gel (10 cm x 10 cm).
NOTE: Loading of prestained molecular weight markers (#59329, 10 µl/lane) to verify electrotransfer and biotinylated protein ladder (#7727, 10 µl/lane) to determine molecular weights are recommended.
NOTE: Volumes are for 10 cm x 10 cm (100 cm2) of membrane; for different sized membranes, adjust volumes accordingly.
* Avoid repeated exposure to skin.
posted June 2005
revised June 2020
Protocol Id: 10
Human, Mouse, Rat, Monkey
Hamster, Xenopus, Zebrafish, Horse, Guinea Pig
Polyclonal antibodies are produced by immunizing animals with a synthetic peptide corresponding to residues surrounding acetylated Lys36 of human histone H3 protein. Antibodies are purified by protein A and peptide affinity chromatography.
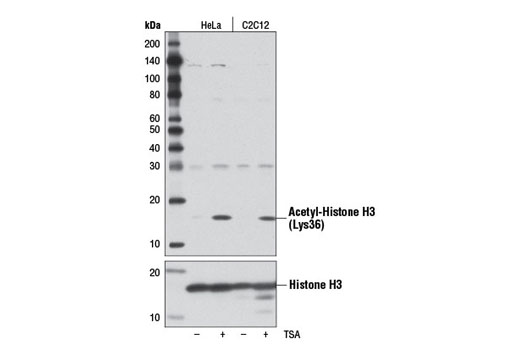
The nucleosome, made up of four core histone proteins (H2A, H2B, H3, and H4), is the primary building block of chromatin. Originally thought to function as a static scaffold for DNA packaging, histones have now been shown to be dynamic proteins, undergoing multiple types of post-translational modifications, including acetylation, phosphorylation, methylation, and ubiquitination (1,2). Histone acetylation occurs mainly on the amino-terminal tail domains of histones H2A (Lys5), H2B (Lys5, 12, 15, and 20), H3 (Lys9, 14, 18, 23, 27, 36, and 56), and H4 (Lys5, 8, 12, and 16) and is important for the regulation of histone deposition, transcriptional activation, DNA replication, recombination, and DNA repair (1-3). Hyper-acetylation of the histone tails neutralizes the positive charge of these domains and is believed to weaken histone-DNA and nucleosome-nucleosome interactions, thereby destabilizing chromatin structure and increasing the accessibility of DNA to various DNA-binding proteins (4,5). In addition, acetylation of specific lysine residues creates docking sites for a protein module called the bromodomain, which binds to acetylated lysine residues (6). Many transcription and chromatin regulatory proteins contain bromodomains and may be recruited to gene promoters, in part, through binding of acetylated histone tails. Histone acetylation is mediated by histone acetyltransferases (HATs), such as CBP/p300, GCN5L2, PCAF, and Tip60, which are recruited to genes by DNA-bound protein factors to facilitate transcriptional activation (3). Deacetylation, which is mediated by histone deacetylases (HDAC and sirtuin proteins), reverses the effects of acetylation and generally facilitates transcriptional repression (7,8).
Histone H3 Lys36 acetylation is mediated by the GCN5 histone acetyltransferase and is conserved in multiple species, from yeast to mammals (9). Acetyl-histone H3 Lys36 is localized predominantly to the promoters of active RNA polymerase II-transcribed genes and overlaps with other acetylation marks associated with transcriptional activation, such as acetyl-histone H3 Lys9 and Lys14 (9). The pattern of acetyl-histone H3 Lys36 is inversely related to that of tri-methyl-histone H3 Lys36, which is found in the gene body of actively transcribed genes.
Explore pathways related to this product.
STRING - Known and Predicted Protein-Protein Interactions.
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.