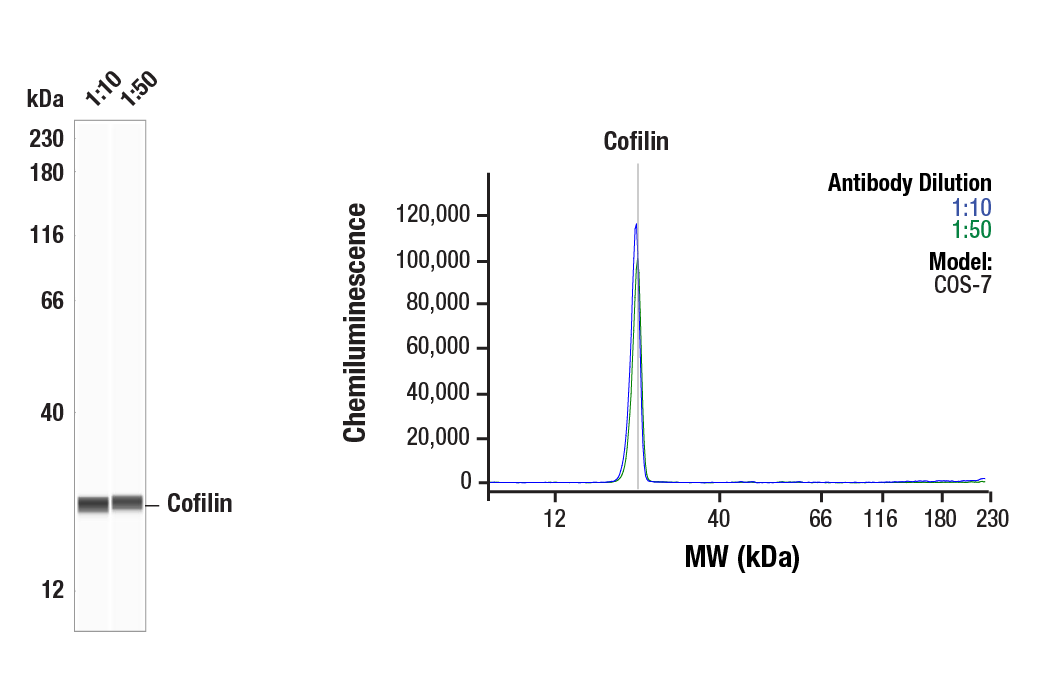
Simple Western™ analysis of lysates (0.1 mg/mL) from Cos-7 cells using Cofilin (D3F9) XP® Rabbit mAb #5175. The virtual lane view (left) shows the target band (as indicated) at 1:10 and 1:50 dilutions of primary antibody. The corresponding electropherogram view (right) plots chemiluminescence by molecular weight along the capillary at 1:10 (blue line) and 1:50 (green line) dilutions of primary antibody. This experiment was performed under reducing conditions on the Jess™ Simple Western instrument from ProteinSimple, a BioTechne brand, using the 12-230 kDa separation module.
| Cat. # | Size | Qty. | Price |
|---|---|---|---|
| 8354T | 1 Kit (7 x 20 microliters) |
|
| Product Includes | Quantity | Applications | Reactivity | MW(kDa) | Isotype |
|---|---|---|---|---|---|
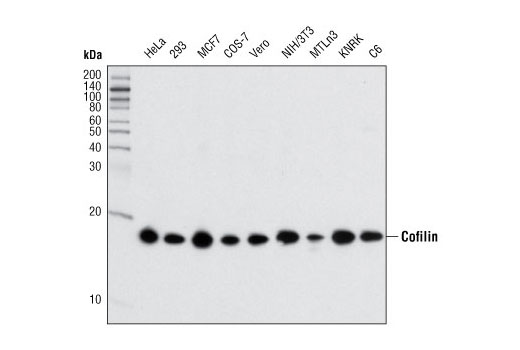
| Cofilin (D3F9) XP® Rabbit mAb 5175 | 20 µl |
|
H M R Mk Dg | 19 | Rabbit IgG |
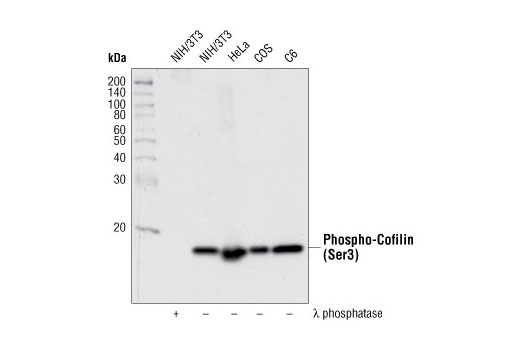
| Phospho-Cofilin (Ser3) (77G2) Rabbit mAb 3313 | 20 µl |
|
H M R Mk B | 19 | Rabbit IgG |
| LIMK2 (8C11) Rabbit mAb 3845 | 20 µl |
|
H Mk | 70 | Rabbit IgG |
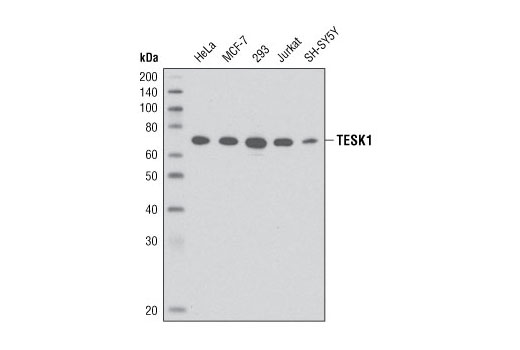
| TESK1 (D49D4) Rabbit mAb 4655 | 20 µl |
|
H | 68 | Rabbit IgG |
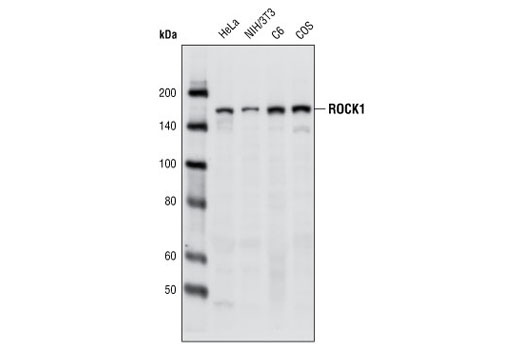
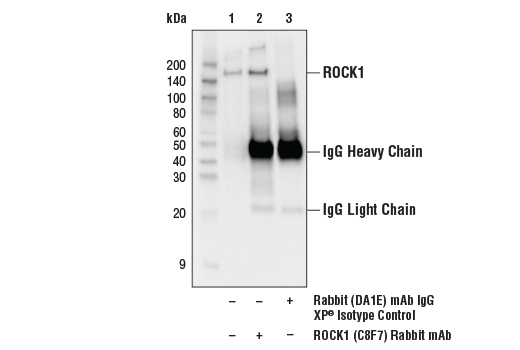
| ROCK1 (C8F7) Rabbit mAb 4035 | 20 µl |
|
H M R Mk | 160 | Rabbit |
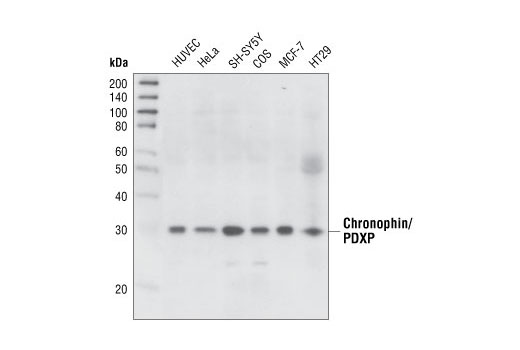
| Chronophin/PDXP (C85E3) Rabbit mAb 4686 | 20 µl |
|
H M R Hm Mk B | 31 | Rabbit IgG |
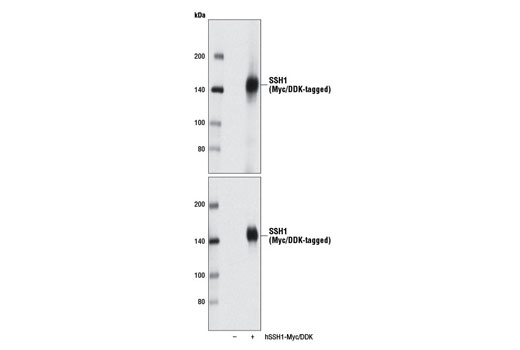
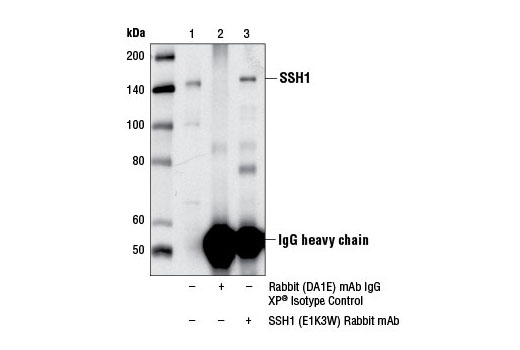
| SSH1 (E1K3W) Rabbit mAb 13578 | 20 µl |
|
H M R | 140 | Rabbit IgG |
| Anti-rabbit IgG, HRP-linked Antibody 7074 | 100 µl |
|
Goat |
Product Information
Monoclonal antibodies are produced by immunizing animals with a synthetic peptide corresponding to central residues of human cofilin1 protein, carboxy-terminal residues of human LIMK2 protein, carboxy-terminal residues of human TESK1 protein, the central sequence of human ROCK1 protein; with recombinant mouse MBP-chronophin protein; with a synthetic phosphopeptide corresponding to residues surrounding Ser3 of human cofilin protein; or with residues surrounding Pro1018 of human SSH1 protein.
Cofilin and actin-depolymerization factor (ADF) are members of a family of essential conserved small actin-binding proteins that play pivotal roles in cytokinesis, endocytosis, embryonic development, stress response, and tissue regeneration (1). In response to stimuli, cofilin promotes the regeneration of actin filaments by severing preexisting filaments (2). The severing activity of cofilin is inhibited by LIMK or TESK phosphorylation at Ser3 of cofilin (3-5). Phosphorylation at Ser3 also regulates cofilin translocation from the nucleus to the cytoplasm (6).
LIM kinases (LIMK1 and LIMK2) are serine/threonine kinases that have two zinc finger motifs, known as LIM motifs, in their amino-terminal regulatory domains (7). LIM kinases are involved in actin cytoskeletal regulation downstream of Rho-family GTPases, PAKs, and ROCK (8,9). PAK1 and ROCK phosphorylate LIMK1 or LIMK2 at the conserved Thr508 or Thr505 residues in the activation loop, increasing LIMK activity (9-11). Activated LIM kinases inhibit the actin depolymerization activity of cofilin by phosphorylation at the amino-terminal Ser3 residue of cofilin (12,13).
Testis-specific kinase 1 (TESK1) is an LIMK-related protein kinase originally identified to be highly expressed in testes and subsequently shown to be expressed in a wide variety of tissues and cell types (14-17). TESK1 phosphorylates the actin severing protein cofilin at Ser3, inactivating cofilin and thus regulating the organization of the actin cytoskeleton (15). Integrin signaling activates TESK1 activity and leads to stress fiber formation and cell spreading (15,18,19). TESK1 is involved in regulation of ERK signaling through its interaction with Spry2 (20) and regulation of cell spreading through its interaction with the focal adhesion protein actopaxin/α-parvin (18).
Chronophin (CIN, PDXP) is a haloacid dehalogenase phosphatase that dephosphorylates cofilin. Alteration of CIN activity through overexpression of either the wildtype or phosphatase-inactive mutant CIN interferes with actin dynamics, cell morphology and cytokinesis (21).
ROCK (Rho-associated kinase), a family of serine/threonine kinases, is an important downstream target of GTPase Rho and plays an important role in Rho-mediated signaling. Two isoforms of ROCK have been identified (ROCK1 and ROCK2). ROCK is composed of N-terminal catalytic, coiled-coil, and C-terminal PH (pleckstrin homology) domains. The C-terminus of ROCK negatively regulates its kinase activity (22,23). Caspase-3-induced cleavage of ROCK1 and direct cleavage of ROCK2 by granzyme B (grB) activates ROCK and leads to phosphorylation of myosin light chain and inhibition of myosin phosphatase (24). This phosphorylation may account for the mechanism by which Rho regulates cytokinesis, cell motility, cell membrane blebbing during apoptosis, and smooth muscle contraction (25-27).
Slingshot homolog 1 (SSH1) can also dephosphorylate LIMK kinases, suppressing LIMK phosphorylation of cofilin (28). In addition, SSH1 modulates actin dynamics by stabilizing F-actin and promoting actin bundling independent of its cofilin phosphatase activity (29). SSH1 activity is regulated by phosphorylation and protein-protein interaction through various signaling pathways (1). Binding of SSH1 to F-actin stimulates its cofilin phosphatase activity (30).
Explore pathways related to this product.
STRING - Known and Predicted Protein-Protein Interactions.
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.