Recombinant antibodies offer several key advantages compared to traditional antibodies. These include superior lot-to-lot consistency, continuous supply, and animal-free manufacturing. As such, recombinant antibodies are seeing increased use for scientific research, especially as a means of addressing the ongoing reproducibility crisis.
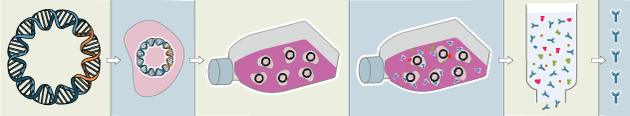
Traditional polyclonal and monoclonal antibodies are the product of normal B cell development and genetic recombination. They are generated by immunizing an animal with an antigen to elicit an immune response. While polyclonal antibodies are secreted by many different B cell clones and recognize multiple antigenic epitopes, monoclonals originate from a single B cell clone and are specific for just one epitope.
Recombinant antibodies are monoclonal, but their production involves in vitro genetic manipulation. After cloning the antibody genes into an expression vector, this is then transfected into an appropriate host cell line for antibody expression. Mammalian cell lines are most commonly used for recombinant antibody production, although cell lines of bacterial, yeast, or insect origin are also suitable.
Because recombinant antibody production involves sequencing the antibody light and heavy chains, it is a highly controlled and reliable process. In contrast, hybridoma-based systems for producing monoclonal antibodies are subject to genetic drift and instability, increasing the potential for lot-to-lot variability or loss of antibody expression. Recombinant antibodies are highly consistent from lot to lot, thereby ensuring reproducible experimental results.
In vitro methods for producing antibodies are amenable to large-scale production, meaning antibody availability is unlikely to become a limiting factor. Moreover, since the recombinant antibody sequence is known, continuity of supply is assured; in situations where an antibody will be used to support large, long-term studies, this can be an especially critical factor.
Unlike traditional methods for antibody production, recombinant approaches avoid the need to use animals. Where polyclonal antibodies are purified directly from the serum of the immunized host, and monoclonals are purified from either hybridoma-derived tissue culture supernatant or ascites, recombinant antibodies are instead purified from the tissue culture supernatants of transfected host cell lines. Regardless of whether an antibody is polyclonal, monoclonal or recombinant, it must always be properly validated in the intended application prior to experimental use. At CST, we adhere to the Hallmarks of Antibody Validation™, six complementary strategies for determining the specificity, sensitivity, and functionality of an antibody in any given assay. By carefully tailoring these strategies to each antibody product, we guarantee that CST antibodies will work as expected, to help you achieve results you can trust.
| Cat. # | Size | Qty. | Price |
|---|---|---|---|
| 14679S | 100 µl |
|
| REACTIVITY | All |
| SENSITIVITY | Endogenous |
| MW (kDa) | |
| Source/Isotype | Rabbit IgG |
Product Information
| Application | Dilution |
|---|---|
| Western Blotting | 1:1000 |
For western blots, incubate membrane with diluted primary antibody in 5% w/v nonfat dry milk, 1X TBS, 0.1% Tween® 20 at 4°C with gentle shaking, overnight.
NOTE: Please refer to primary antibody product webpage for recommended antibody dilution.
NOTE: Prepare solutions with reverse osmosis deionized (RODI) or equivalent grade water.
Load 20 µl onto SDS-PAGE gel (10 cm x 10 cm).
NOTE: Loading of prestained molecular weight markers (#59329, 10 µl/lane) to verify electrotransfer and biotinylated protein ladder (#7727, 10 µl/lane) to determine molecular weights are recommended.
NOTE: Volumes are for 10 cm x 10 cm (100 cm2) of membrane; for different sized membranes, adjust volumes accordingly.
* Avoid repeated exposure to skin.
posted June 2005
revised June 2020
Protocol Id: 263
All Species Expected
All Species Expected
MultiMab® rabbit monoclonal antibody mixes are prepared by combining individual rabbit monoclonal clones in optimized ratios for the approved applications. Each antibody in the mix is carefully selected based on motif recognition and performance in multiple assays. Each mix is engineered to yield the broadest possible coverage of the modification being studied while ensuring a high degree of specificity for the modification or motif.
Methylation of lysine residues is a common regulatory post-translational modification (PTM) that results in the mono-, di-, or tri-methylation of lysine at ε-amine groups by protein lysine methyltransferases (PKMTs). Two PKMT groups are recognized based on structure and catalytic mechanism: class I methyltransferases or seven β strand enzymes, and SET domain-containing class V methyltransferases. Both use the methyl donor S-adenosyl-L-methionine to methylate histone and non-histone proteins. Class I methyltransferases methylate amino acids, DNA, and RNA (1,2). Six methyl-lysine-interacting protein families are distinguished based on binding domains: MBT, PHD finger, Tudor, PWWP, WD40 repeat, and chromodomains. Many of these display differential binding preferences based on lysine methylation state (3). KDM1 subfamily lysine demethylases catalyze demethylation of mono- and di-methyl lysines, while 2-oxoglutarate-dependent JmjC (KDM2-7) subfamily enzymes also modify tri-methyl lysine residues (4).Most PKMT substrates are histone proteins and transcription factors, emphasizing the importance of lysine methylation in regulating chromatin structure and gene expression. Lys9 of histone H3 is mono- or di-methylated by G9A/GLP and tri-methylated by SETDB1 to activate transcription. JHDM3A-mediated demethylation of the same residue creates mono-methyl Lys9 and inhibits gene transcription (5). Tumor suppressor p53 is regulated by methylation of at least four sites. p53-mediated transcription is repressed following mono-methylation of p53 at Lys370 by SMYD2; di-methylation at the same residue further inhibits p53 by preventing association with 53BP1. Concomitant di-methylation at Lys382 inhibits p53 ubiquitination following DNA damage. Mono-methylation at Lys382 by SET8 suppresses p53 transcriptional activity, while SET7/9 mono-methylation at Lys372 inhibits SMYD2 methylation at Lys370 and stabilizes the p53 protein. Di-methylation at Lys373 by G9A/GLP inhibits p53-mediated apoptosis and correlates with tri-methylation of histone H3 Lys9 at the p21 promoter (1,6). Overexpression of PKMTs is associated with multiple forms of human cancer, which has generated tremendous interest in targeting protein lysine methyltransferases in drug discovery research.
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.
![Western Blotting Image 1: Mono-Methyl Lysine [mme-K] MultiMab® Rabbit mAb mix](http://media.cellsignal.com/product/image/14679_fig01_W_263_20210423082341.jpg)
