| REACTIVITY | H |
Product Information
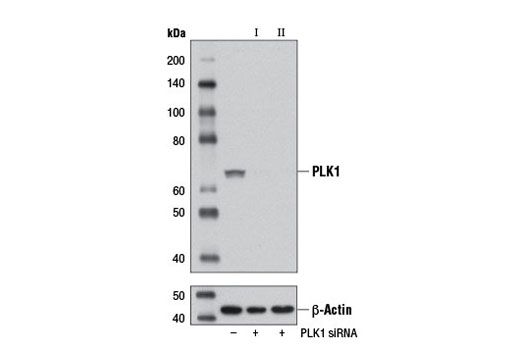
CST recommends transfection with 100 nM SignalSilence® PLK1 siRNA I 48 to 72 hours prior to cell lysis. For transfection procedure, follow protocol provided by the transfection reagent manufacturer. Please feel free to contact CST with any questions on use.
Each vial contains the equivalent of 100 transfections, which corresponds to a final siRNA concentration of 100 nM per transfection in a 24-well plate with a total volume of 300 μl per well.
Oligonucleotide synthesis is monitored base by base through trityl analysis to ensure appropriate coupling efficiency. The oligo is subsequently purified by affinity-solid phase extraction. The annealed RNA duplex is further analyzed by mass spectrometry to verify the exact composition of the duplex. Each lot is compared to the previous lot by mass spectrometry to ensure maximum lot-to-lot consistency.
At least four distinct polo-like kinases exist in mammalian cells: PLK1, PLK2, PLK3, and PLK4/SAK (1). PLK1 apparently plays many roles during mitosis, particularly in regulating mitotic entry and exit. The mitosis promoting factor (MPF), cdc2/cyclin B1, is activated by dephosphorylation of cdc2 (Thr14/Tyr15) by cdc25C. PLK1 phosphorylates cdc25C at Ser198 and cyclin B1 at Ser133, causing translocation of these proteins from the cytoplasm to the nucleus (2-5). PLK1 phosphorylation of Myt1 at Ser426 and Thr495 has been proposed to inactivate Myt1, one of the kinases known to phosphorylate cdc2 at Thr14/Tyr15 (6). Polo-like kinases also phosphorylate the cohesin subunit SCC1, causing cohesin displacement from chromosome arms that allow for proper cohesin localization to centromeres (7). Mitotic exit requires activation of the anaphase promoting complex (APC) (8), a ubiquitin ligase responsible for removal of cohesin at centromeres, and degradation of securin, cyclin A, cyclin B1, Aurora A, and cdc20 (9). PLK1 phosphorylation of the APC subunits Apc1, cdc16, and cdc27 has been demonstrated in vitro and has been proposed as a mechanism by which mitotic exit is regulated (10,11).
Substitution of Thr210 with Asp has been reported to elevate PLK1 kinase activity and delay/arrest cells in mitosis, while a Ser137Asp substitution leads to S-phase arrest (12). In addition, while DNA damage has been found to inhibit PLK1 kinase activity, the Thr210Asp mutant is resistant to this inhibition (13). PLK1 has been reported to be phosphorylated in vivo at Ser137 and Thr210 in mitosis; DNA damage prevents phosphorylation at these sites (14).
Explore pathways related to this product.
STRING - Known and Predicted Protein-Protein Interactions.
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.
| Cat. # | Size | Qty. | Price |
|---|---|---|---|
| 6292S | 300 µl (3 nmol) |
|